

Chemotaxis Control of Transient Cell Aggregation
- PMID: 26216846
- PMCID: PMC4573731
- DOI: 10.1128/JB.00121-15
Chemotaxis Control of Transient Cell Aggregation
Abstract
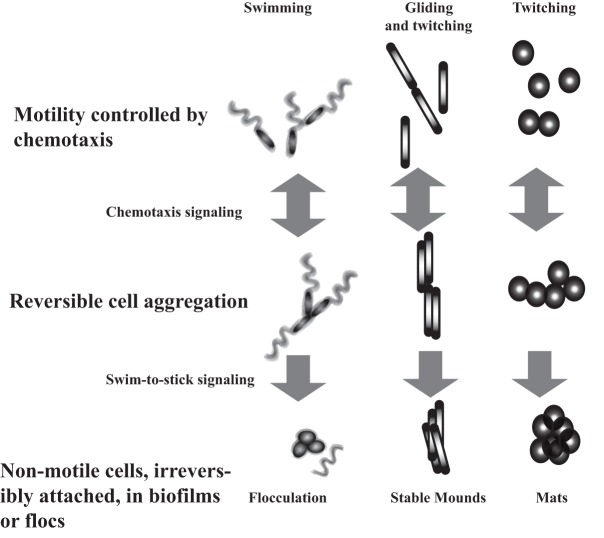
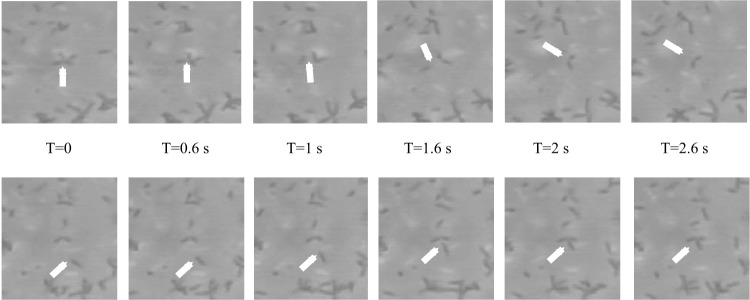
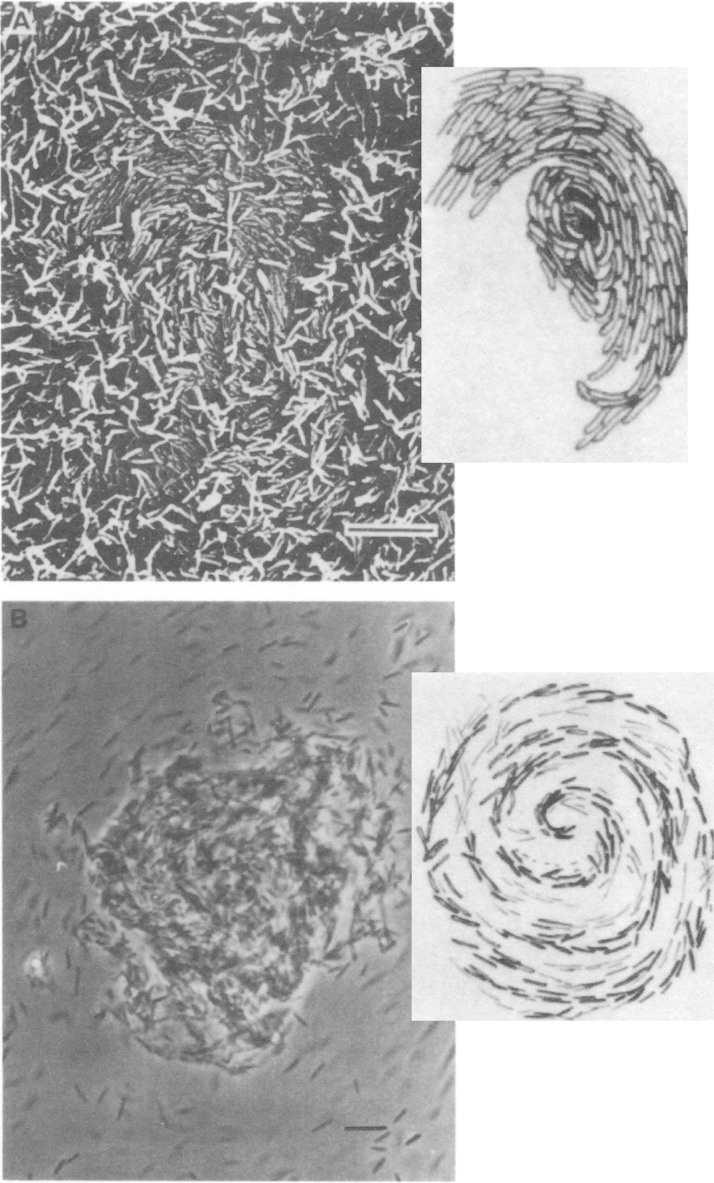
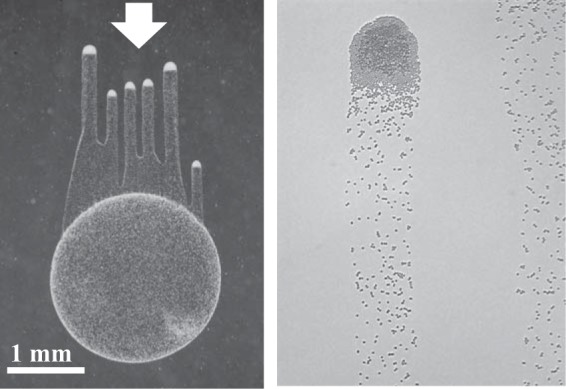
Chemotaxis affords motile cells the ability to rapidly respond to environmental challenges by navigating cells to niches favoring growth. Such a property results from the activities of dedicated signal transduction systems on the motility apparatus, such as flagella, type IV pili, and gliding machineries. Once cells have reached a niche with favorable conditions, they often stop moving and aggregate into complex communities termed biofilms. An intermediate and reversible stage that precedes commitment to permanent adhesion often includes transient cell-cell contacts between motile cells. Chemotaxis signaling has been implicated in modulating the transient aggregation of motile cells. Evidence further indicates that chemotaxis-dependent transient cell aggregation events are behavioral responses to changes in metabolic cues that temporarily prohibit permanent attachment by maintaining motility and chemotaxis. This minireview discusses a few examples illustrating the role of chemotaxis signaling in the initiation of cell-cell contacts in bacteria moving via flagella, pili, or gliding.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Flemming H-C, Wingender J. 2010. The biofilm matrix. Nat Rev Microbiol 8:623–633. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
