

Cancer as a mitochondrial metabolic disease
- PMID: 26217661
- PMCID: PMC4493566
- DOI: 10.3389/fcell.2015.00043
Cancer as a mitochondrial metabolic disease
Abstract
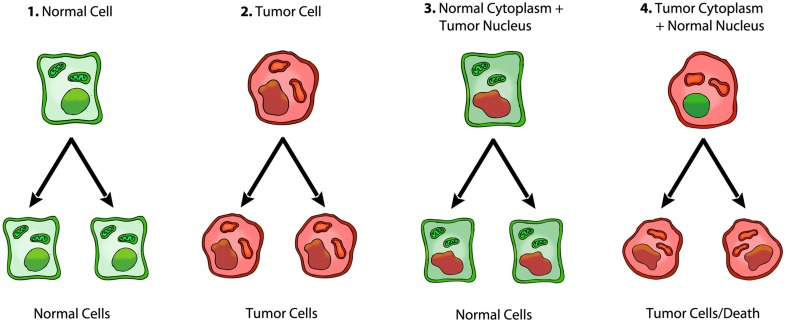
Cancer is widely considered a genetic disease involving nuclear mutations in oncogenes and tumor suppressor genes. This view persists despite the numerous inconsistencies associated with the somatic mutation theory. In contrast to the somatic mutation theory, emerging evidence suggests that cancer is a mitochondrial metabolic disease, according to the original theory of Otto Warburg. The findings are reviewed from nuclear cytoplasm transfer experiments that relate to the origin of cancer. The evidence from these experiments is difficult to reconcile with the somatic mutation theory, but is consistent with the notion that cancer is primarily a mitochondrial metabolic disease.
Keywords: Warburg effect; carcinogenesis; cybrids; fermentation; microenvironment; mitochondria; oxidative phosphorylation; tumorigenesis.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources