

Human Enterovirus Nonstructural Protein 2CATPase Functions as Both an RNA Helicase and ATP-Independent RNA Chaperone
- PMID: 26218680
- PMCID: PMC4517893
- DOI: 10.1371/journal.ppat.1005067
Human Enterovirus Nonstructural Protein 2CATPase Functions as Both an RNA Helicase and ATP-Independent RNA Chaperone
Abstract
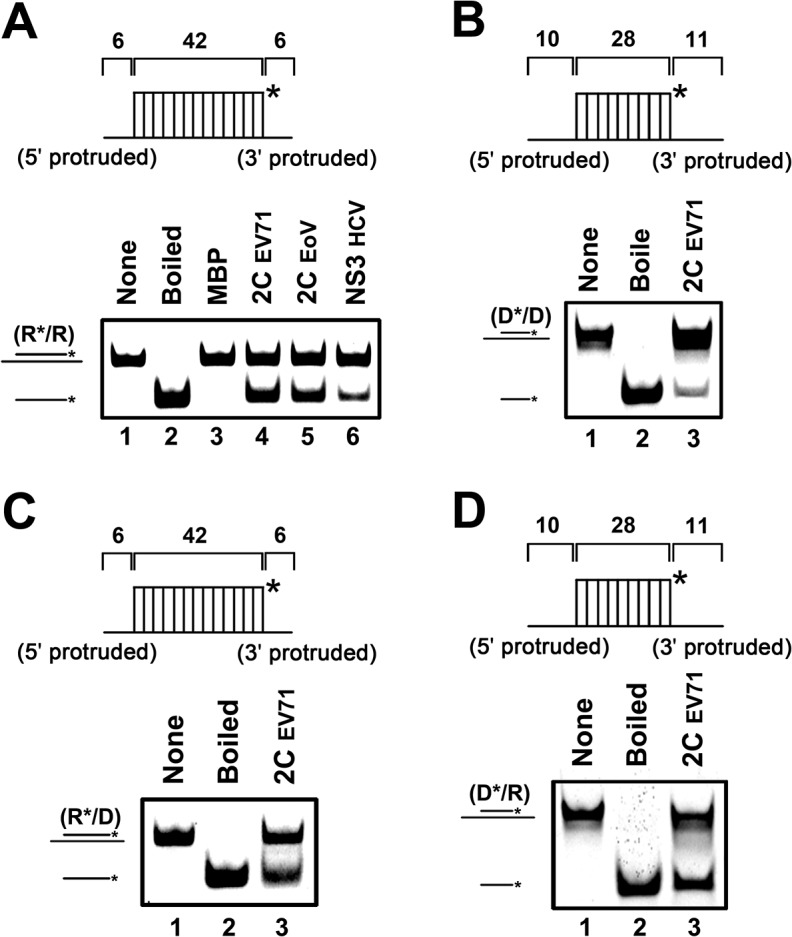
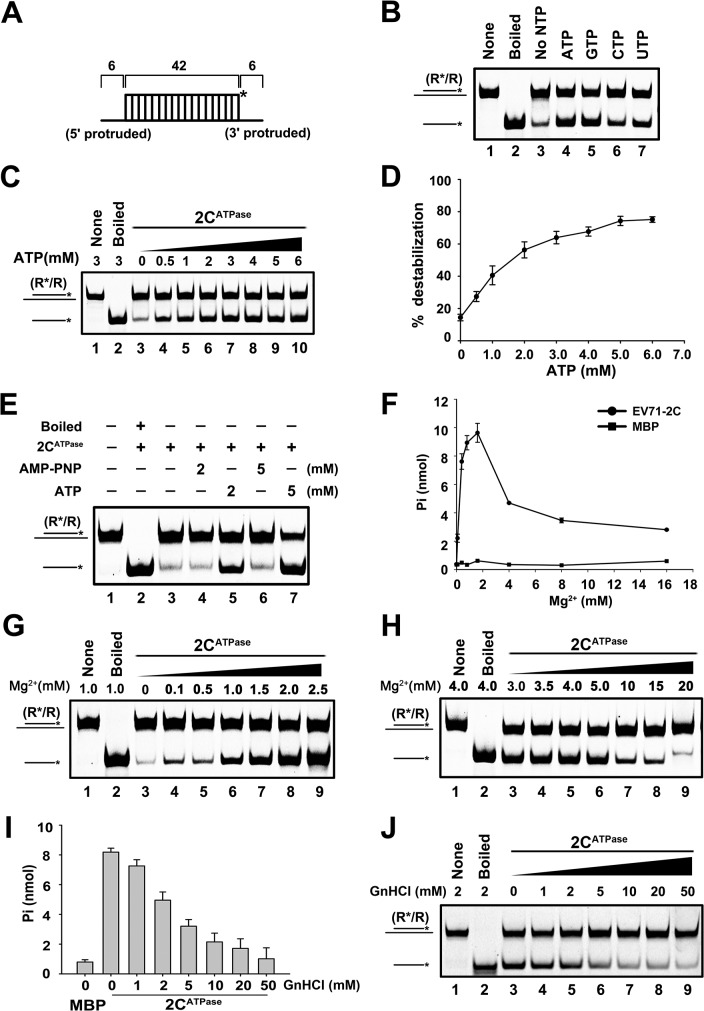
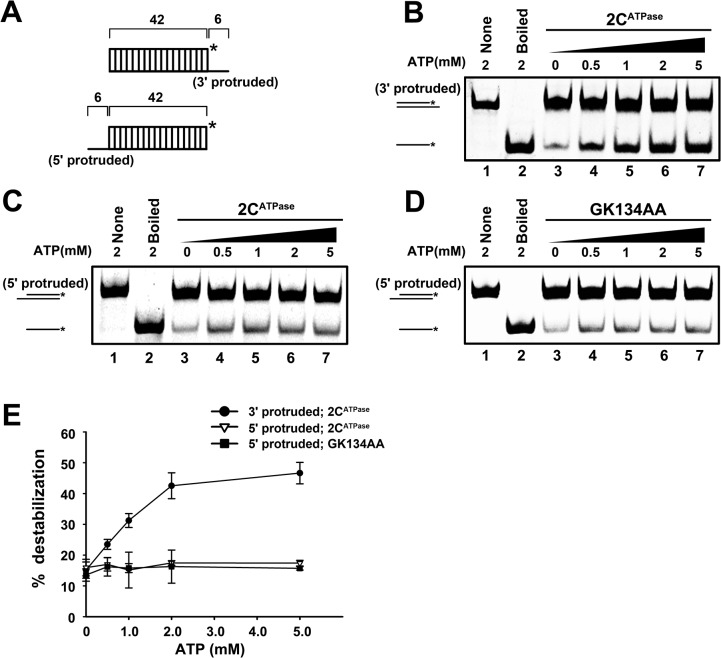
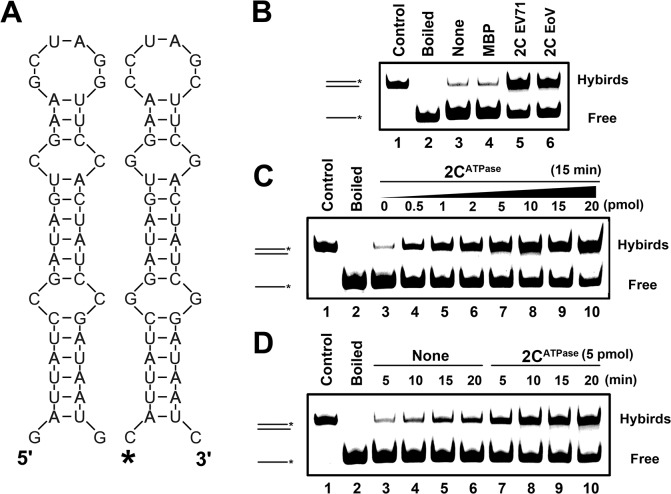
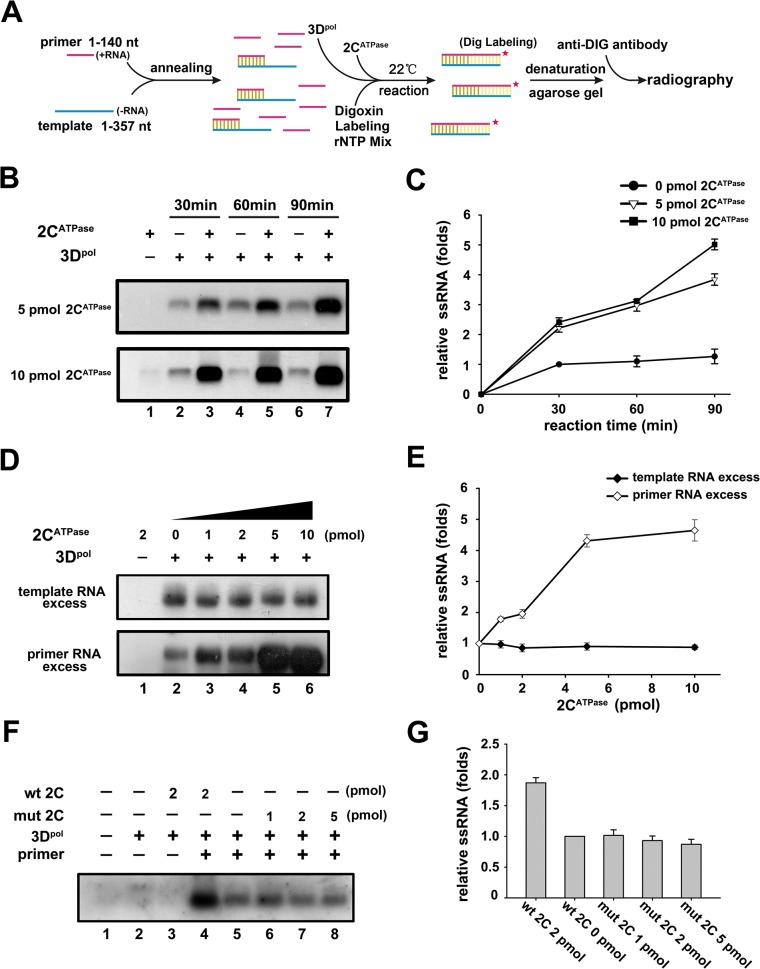
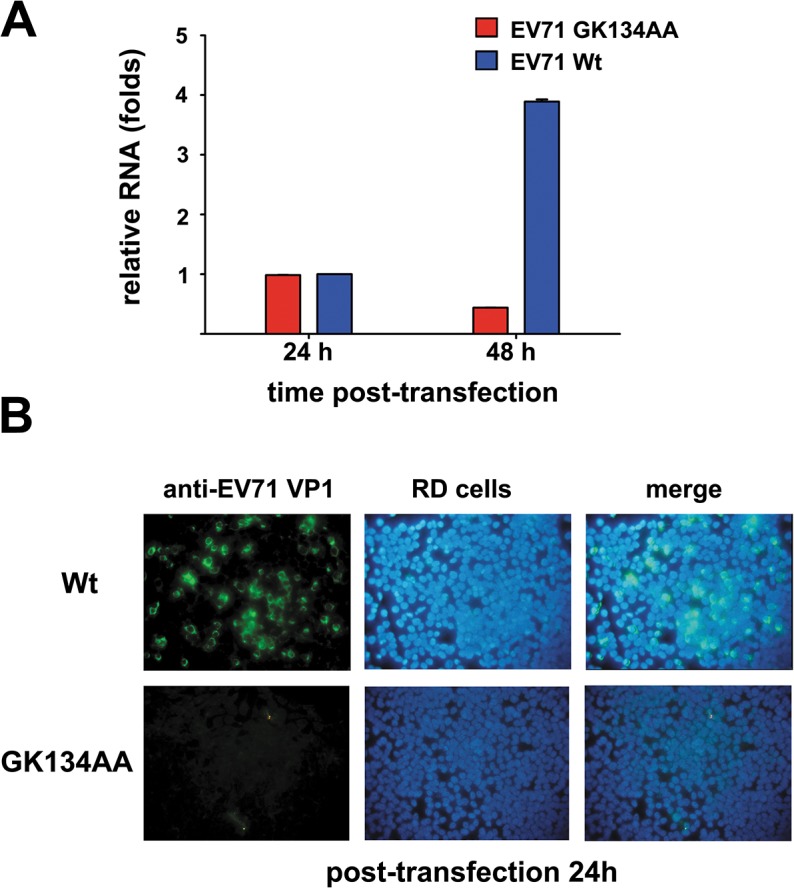
RNA helicases and chaperones are the two major classes of RNA remodeling proteins, which function to remodel RNA structures and/or RNA-protein interactions, and are required for all aspects of RNA metabolism. Although some virus-encoded RNA helicases/chaperones have been predicted or identified, their RNA remodeling activities in vitro and functions in the viral life cycle remain largely elusive. Enteroviruses are a large group of positive-stranded RNA viruses in the Picornaviridae family, which includes numerous important human pathogens. Herein, we report that the nonstructural protein 2CATPase of enterovirus 71 (EV71), which is the major causative pathogen of hand-foot-and-mouth disease and has been regarded as the most important neurotropic enterovirus after poliovirus eradication, functions not only as an RNA helicase that 3'-to-5' unwinds RNA helices in an adenosine triphosphate (ATP)-dependent manner, but also as an RNA chaperone that destabilizes helices bidirectionally and facilitates strand annealing and complex RNA structure formation independently of ATP. We also determined that the helicase activity is based on the EV71 2CATPase middle domain, whereas the C-terminus is indispensable for its RNA chaperoning activity. By promoting RNA template recycling, 2CATPase facilitated EV71 RNA synthesis in vitro; when 2CATPase helicase activity was impaired, EV71 RNA replication and virion production were mostly abolished in cells, indicating that 2CATPase-mediated RNA remodeling plays a critical role in the enteroviral life cycle. Furthermore, the RNA helicase and chaperoning activities of 2CATPase are also conserved in coxsackie A virus 16 (CAV16), another important enterovirus. Altogether, our findings are the first to demonstrate the RNA helicase and chaperoning activities associated with enterovirus 2CATPase, and our study provides both in vitro and cellular evidence for their potential roles during viral RNA replication. These findings increase our understanding of enteroviruses and the two types of RNA remodeling activities.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
