

Dengue E Protein Domain III-Based DNA Immunisation Induces Strong Antibody Responses to All Four Viral Serotypes
- PMID: 26218926
- PMCID: PMC4517776
- DOI: 10.1371/journal.pntd.0003947
Dengue E Protein Domain III-Based DNA Immunisation Induces Strong Antibody Responses to All Four Viral Serotypes
Abstract
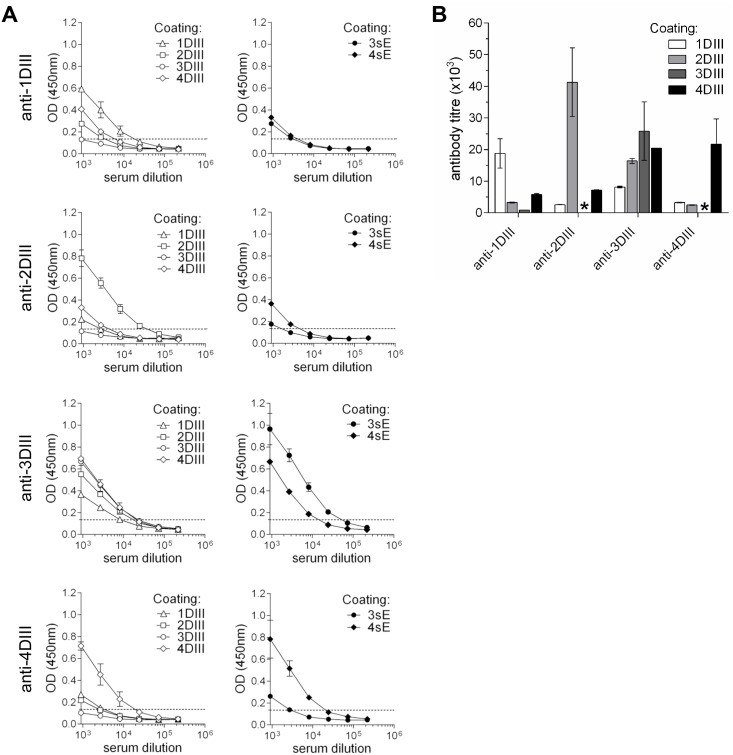
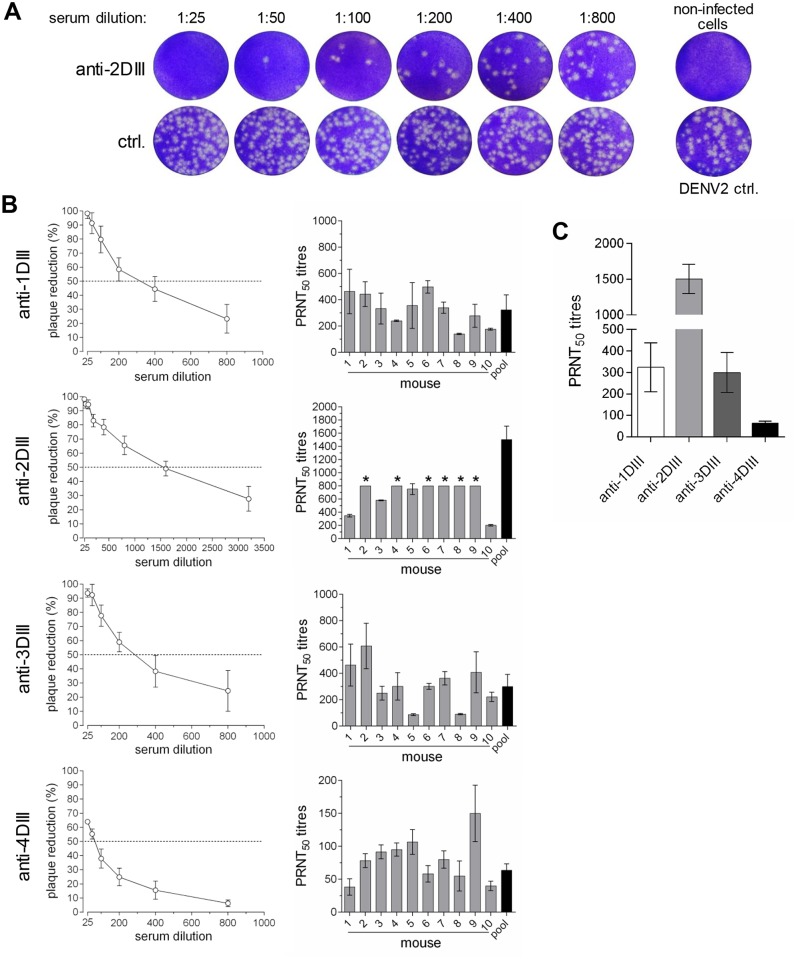
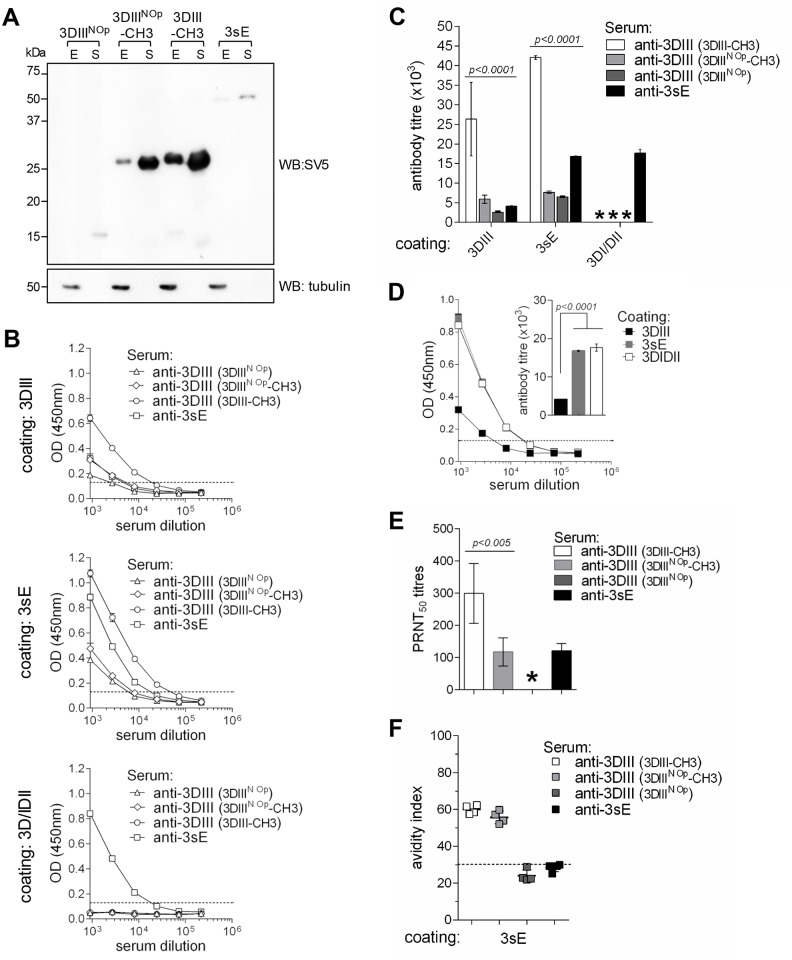
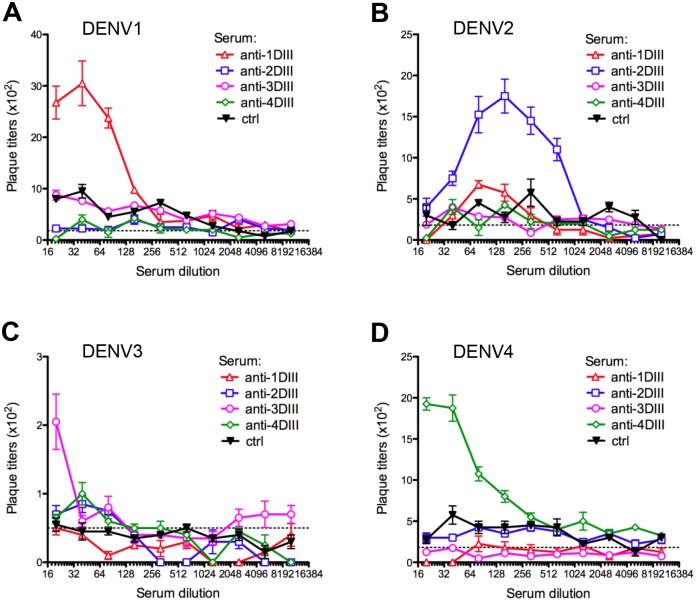
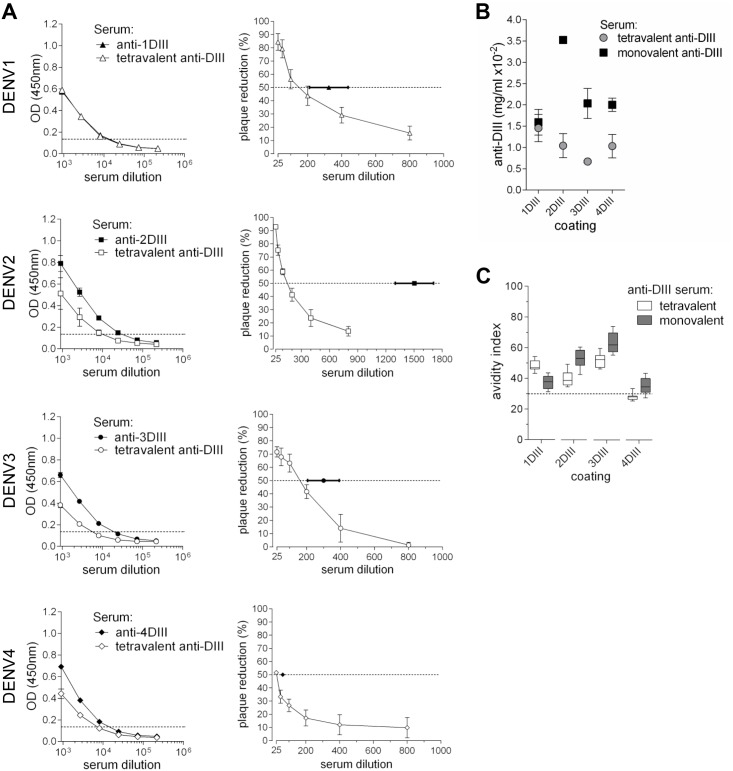
Dengue virus (DENV) infection is a major emerging disease widely distributed throughout the tropical and subtropical regions of the world affecting several millions of people. Despite constants efforts, no specific treatment or effective vaccine is yet available. Here we show a novel design of a DNA immunisation strategy that resulted in the induction of strong antibody responses with high neutralisation titres in mice against all four viral serotypes. The immunogenic molecule is an engineered version of the domain III (DIII) of the virus E protein fused to the dimerising CH3 domain of the IgG immunoglobulin H chain. The DIII sequences were also codon-optimised for expression in mammalian cells. While DIII alone is very poorly secreted, the codon-optimised fusion protein is rightly expressed, folded and secreted at high levels, thus inducing strong antibody responses. Mice were immunised using gene-gun technology, an efficient way of intradermal delivery of the plasmid DNA, and the vaccine was able to induce neutralising titres against all serotypes. Additionally, all sera showed reactivity to a recombinant DIII version and the recombinant E protein produced and secreted from mammalian cells in a mono-biotinylated form when tested in a conformational ELISA. Sera were also highly reactive to infective viral particles in a virus-capture ELISA and specific for each serotype as revealed by the low cross-reactive and cross-neutralising activities. The serotype specific sera did not induce antibody dependent enhancement of infection (ADE) in non-homologous virus serotypes. A tetravalent immunisation protocol in mice showed induction of neutralising antibodies against all four dengue serotypes as well.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: A patent on the invention of the antigen construct for DNA vaccination has been filed by MP, MB, and ORB. This does not alter our adherence to all PLOS policies on sharing data and materials.
Figures
Similar articles
-
Long-term stability of antibody responses elicited by Dengue virus envelope DIII-based DNA vaccines.J Gen Virol. 2018 Aug;99(8):1078-1085. doi: 10.1099/jgv.0.001094. Epub 2018 Jun 20. J Gen Virol. 2018. PMID: 29923817
-
An Envelope-Modified Tetravalent Dengue Virus-Like-Particle Vaccine Has Implications for Flavivirus Vaccine Design.J Virol. 2017 Nov 14;91(23):e01181-17. doi: 10.1128/JVI.01181-17. Print 2017 Dec 1. J Virol. 2017. PMID: 28956764 Free PMC article.
-
Secretion of dengue virus envelope protein ectodomain from mammalian cells is dependent on domain II serotype and affects the immune response upon DNA vaccination.J Gen Virol. 2015 Nov;96(11):3265-3279. doi: 10.1099/jgv.0.000278. Epub 2015 Sep 9. J Gen Virol. 2015. PMID: 26358704
-
Recent progress in dengue vaccine research and development.Curr Opin Mol Ther. 2010 Feb;12(1):31-8. Curr Opin Mol Ther. 2010. PMID: 20140814 Review.
-
A recombinant, chimeric tetravalent dengue vaccine candidate based on a dengue virus serotype 2 backbone.Expert Rev Vaccines. 2016;15(4):497-508. doi: 10.1586/14760584.2016.1128328. Epub 2016 Feb 22. Expert Rev Vaccines. 2016. PMID: 26635182 Review.
Cited by
-
Adjuvant-Mediated Epitope Specificity and Enhanced Neutralizing Activity of Antibodies Targeting Dengue Virus Envelope Protein.Front Immunol. 2017 Sep 25;8:1175. doi: 10.3389/fimmu.2017.01175. eCollection 2017. Front Immunol. 2017. PMID: 28993770 Free PMC article.
-
Adaptive Immunity to Dengue Virus: Slippery Slope or Solid Ground for Rational Vaccine Design?Pathogens. 2020 Jun 15;9(6):470. doi: 10.3390/pathogens9060470. Pathogens. 2020. PMID: 32549226 Free PMC article. Review.
-
DENV2 Pseudoviral Particles with Unprocessed Capsid Protein Are Assembled and Infectious.Viruses. 2019 Dec 25;12(1):27. doi: 10.3390/v12010027. Viruses. 2019. PMID: 31881703 Free PMC article.
-
Dendritic Cell Targeting Using a DNA Vaccine Induces Specific Antibodies and CD4+ T Cells to the Dengue Virus Envelope Protein Domain III.Front Immunol. 2019 Jan 29;10:59. doi: 10.3389/fimmu.2019.00059. eCollection 2019. Front Immunol. 2019. PMID: 30761131 Free PMC article.
-
Development of MVA-d34 Tetravalent Dengue Vaccine: Design and Immunogenicity.Vaccines (Basel). 2023 Apr 12;11(4):831. doi: 10.3390/vaccines11040831. Vaccines (Basel). 2023. PMID: 37112743 Free PMC article.
References
-
- WHO. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control Geneva: World Health Organization Press; 2009 [cited 2014 November 13]. http://www.who.int/tdr/publications/documents/dengue-diagnosis.pdf. - PubMed
-
- Gulland A. Burden of dengue fever is higher than previously thought. BMJ (Clinical research ed). 2013;347:f6280. Epub 2013/10/18. - PubMed
-
- WHO. Global Strategy for Dengue Prevention and Control, 2012–2020. Geneva: World Health Organization Press; 2012 [cited 2014 November 13]. http://reliefweb.int/sites/reliefweb.int/files/resources/9789241504034_e....
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
