

FANCD2-associated nuclease 1, but not exonuclease 1 or flap endonuclease 1, is able to unhook DNA interstrand cross-links in vitro
- PMID: 26221031
- PMCID: PMC4566234
- DOI: 10.1074/jbc.M115.663666
FANCD2-associated nuclease 1, but not exonuclease 1 or flap endonuclease 1, is able to unhook DNA interstrand cross-links in vitro
Abstract
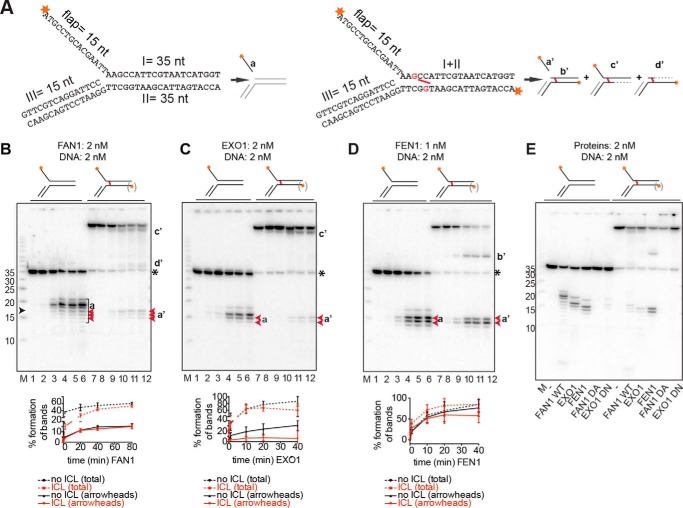
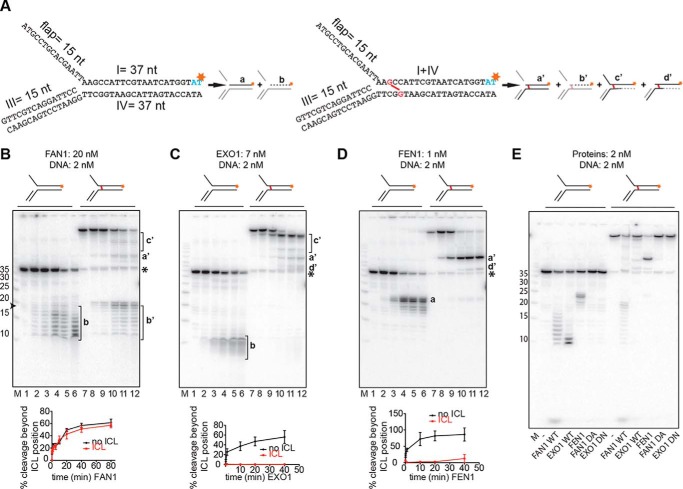
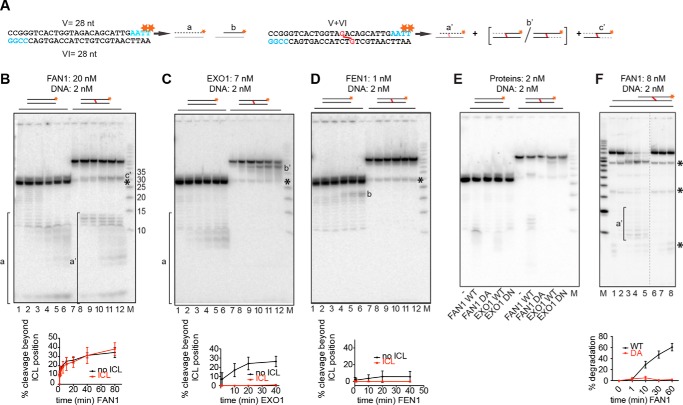
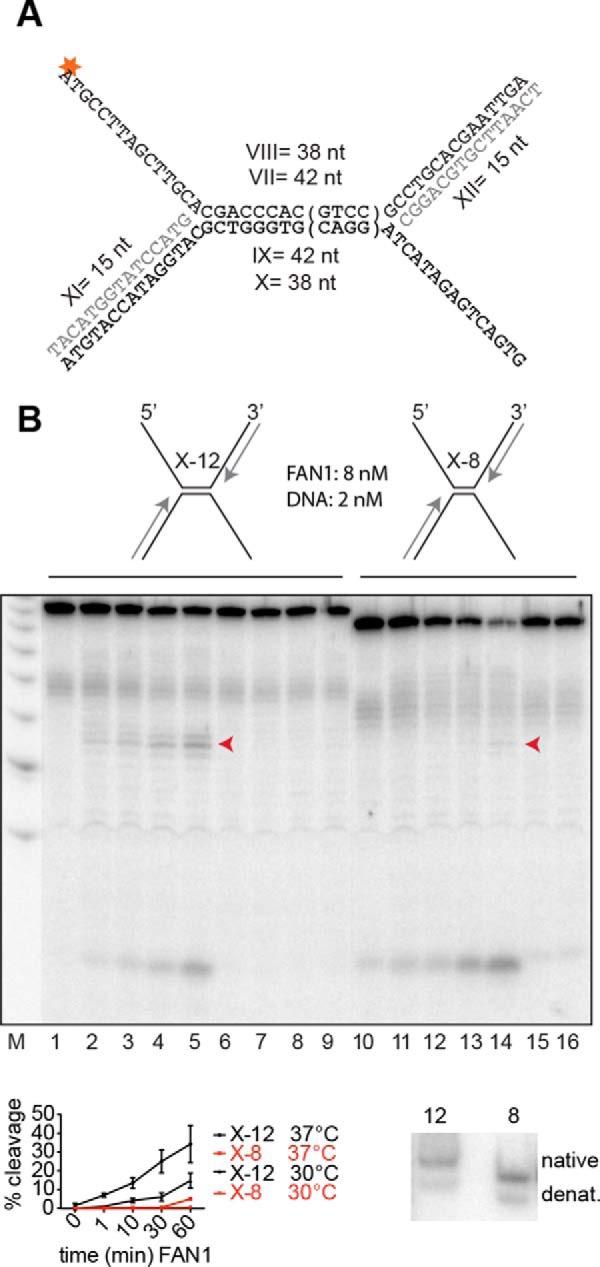
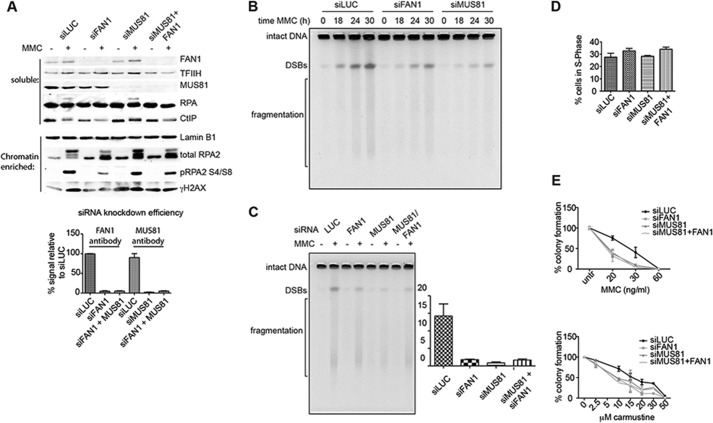
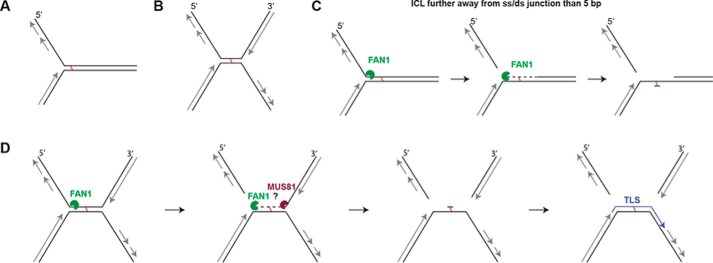
Cisplatin and its derivatives, nitrogen mustards and mitomycin C, are used widely in cancer chemotherapy. Their efficacy is linked primarily to their ability to generate DNA interstrand cross-links (ICLs), which effectively block the progression of transcription and replication machineries. Release of this block, referred to as unhooking, has been postulated to require endonucleases that incise one strand of the duplex on either side of the ICL. Here we investigated how the 5' flap nucleases FANCD2-associated nuclease 1 (FAN1), exonuclease 1 (EXO1), and flap endonuclease 1 (FEN1) process a substrate reminiscent of a replication fork arrested at an ICL. We now show that EXO1 and FEN1 cleaved the substrate at the boundary between the single-stranded 5' flap and the duplex, whereas FAN1 incised it three to four nucleotides in the double-stranded region. This affected the outcome of processing of a substrate containing a nitrogen mustard-like ICL two nucleotides in the duplex region because FAN1, unlike EXO1 and FEN1, incised the substrate predominantly beyond the ICL and, therefore, failed to release the 5' flap. We also show that FAN1 was able to degrade a linear ICL substrate. This ability of FAN1 to traverse ICLs in DNA could help to elucidate its biological function, which is currently unknown.
Keywords: DNA damage; DNA endonuclease; DNA repair; DNA-protein interaction; enzyme mechanism; exonuclease; interstrand cross-links.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Schärer O. D. (2005) DNA interstrand crosslinks: natural and drug-induced DNA adducts that induce unique cellular responses. ChemBioChem 6, 27–32 - PubMed
-
- Hinz J. M. (2010) Role of homologous recombination in DNA interstrand crosslink repair. Environ. Mol. Mutagen. 51, 582–603 - PubMed
-
- Crossan G. P., Patel K. J. (2012) The Fanconi anaemia pathway orchestrates incisions at sites of crosslinked DNA. J. Pathol. 226, 326–337 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
