

Viral gene expression potentiates reovirus-induced necrosis
- PMID: 26226583
- PMCID: PMC4567420
- DOI: 10.1016/j.virol.2015.06.018
Viral gene expression potentiates reovirus-induced necrosis
Abstract
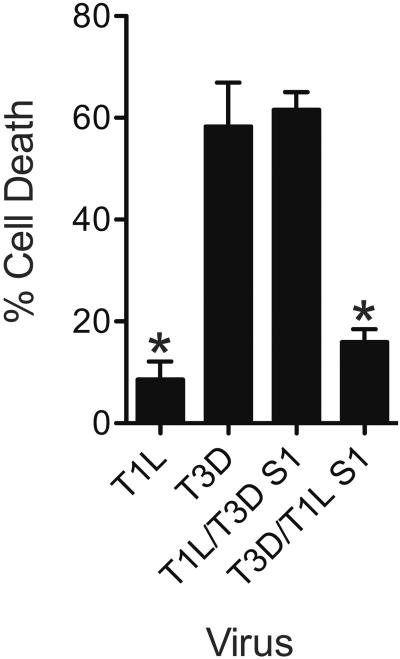
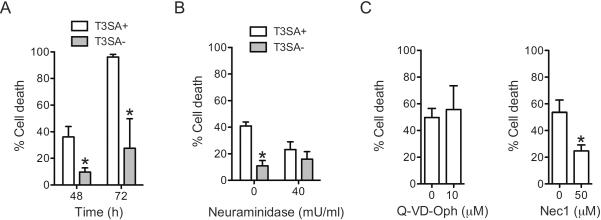
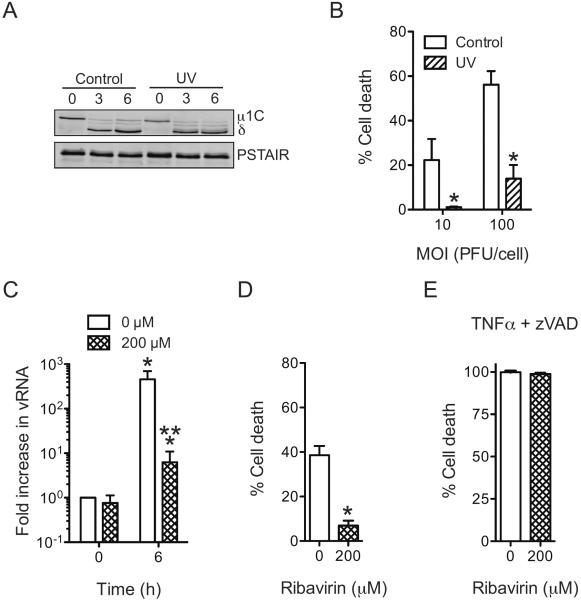
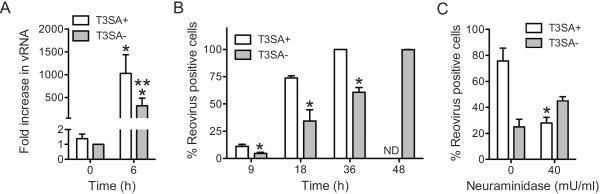
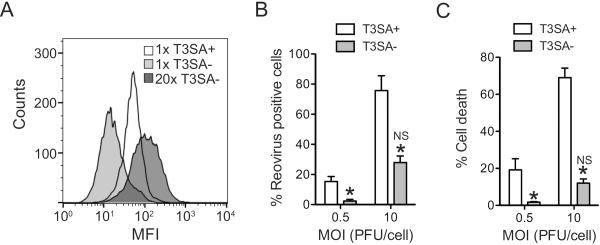
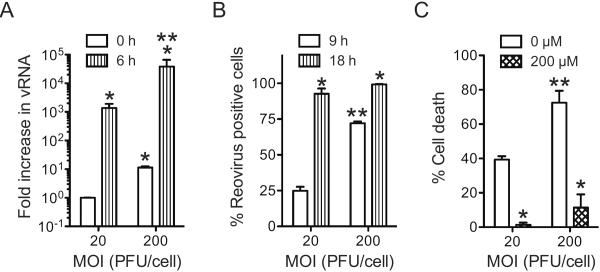
Infection of some cell types by reovirus evokes a caspase-independent form of cell death resembling necrosis. While reovirus strain T3D induces necrosis much more efficiently than strain T1L, which viral components contribute to this difference is not known. In this study, we identified that the sialic acid binding property of the reovirus σ1 protein affects necrosis efficiency. We found that in addition to sialic acid engagement by the virus particles, viral gene expression, in the form of viral RNA or protein synthesis, is also required for necrosis induction. Our studies reveal that sialic acid does not directly participate in necrosis induction by initiating a signaling pathway. Instead, sialic acid engagement augments necrosis induction indirectly, by increasing reovirus gene expression in each infected cell. Comparison of our results with previous studies suggests that reovirus-induced apoptosis and necrosis are initiated by distinct stages of viral infection.
Keywords: Cell death; Necrosis; Reovirus.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Barton ES, Connolly JL, Forrest JC, Chappell JD, Dermody TS. Utilization of sialic acid as a coreceptor enhances reovirus attachment by multistep adhesion strengthening. J Biol Chem. 2001a;276:2200–2211. - PubMed
-
- Barton ES, Forrest JC, Connolly JL, Chappell JD, Liu Y, Schnell FJ, Nusrat A, Parkos CA, Dermody TS. Junction adhesion molecule is a receptor for reovirus. Cell. 2001b;104:441–451. - PubMed
-
- Berard A, Coombs KM. Mammalian reoviruses: propagation, quantification, and storage. Current protocols in microbiology. 2009;Chapter 15(Unit15C):11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
