

How genetic errors in GPCRs affect their function: Possible therapeutic strategies
- PMID: 26229975
- PMCID: PMC4516391
- DOI: 10.1016/j.gendis.2015.02.005
How genetic errors in GPCRs affect their function: Possible therapeutic strategies
Abstract
Activating and inactivating mutations in numerous human G protein-coupled receptors (GPCRs) are associated with a wide range of disease phenotypes. Here we use several class A GPCRs with a particularly large set of identified disease-associated mutations, many of which were biochemically characterized, along with known GPCR structures and current models of GPCR activation, to understand the molecular mechanisms yielding pathological phenotypes. Based on this mechanistic understanding we also propose different therapeutic approaches, both conventional, using small molecule ligands, and novel, involving gene therapy.
Keywords: Activation; Agonist; GPCR; Gene therapy; Genetic disorder; Mutation.
Figures
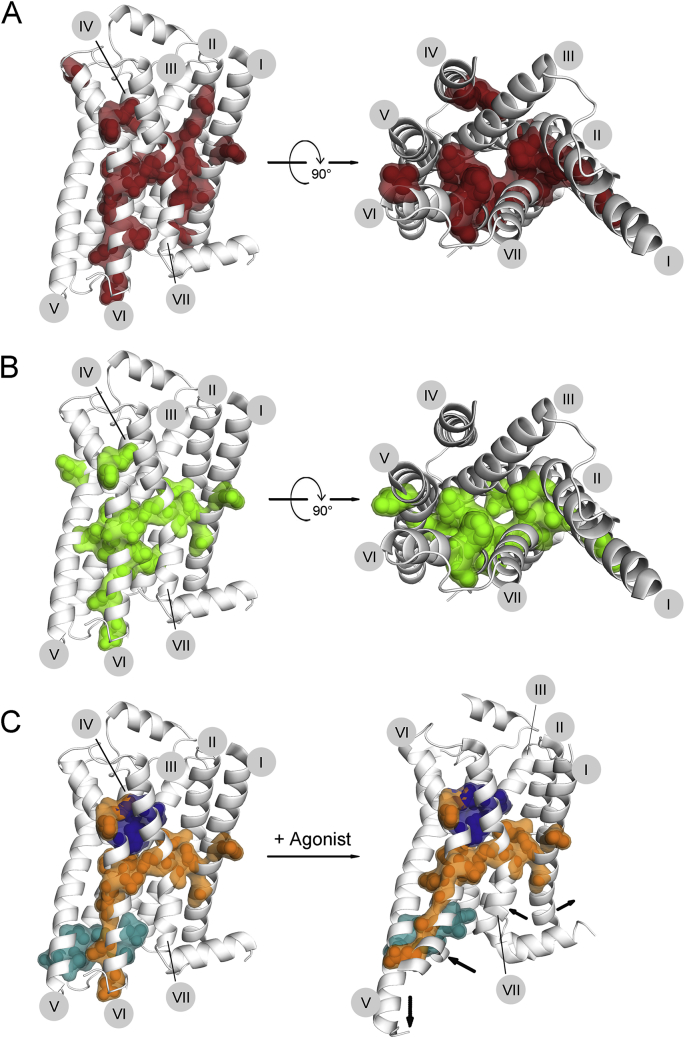
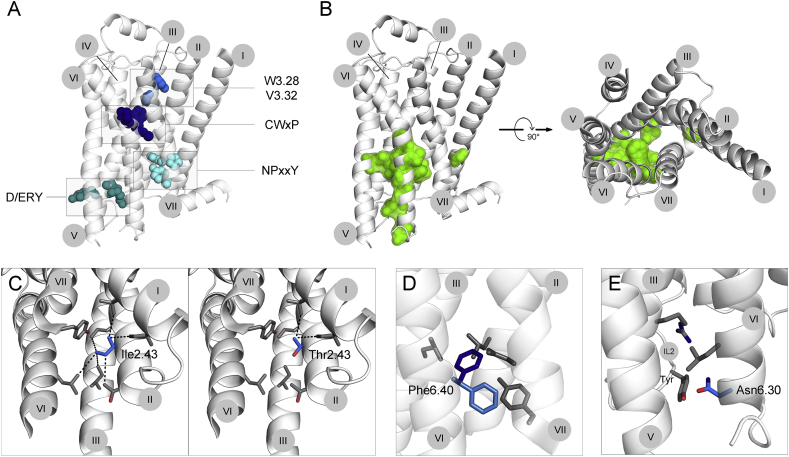
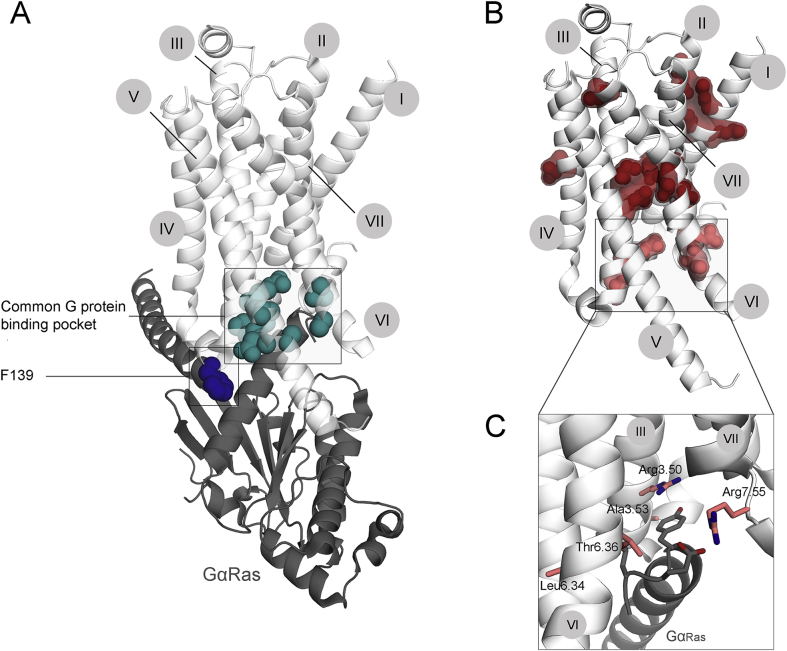
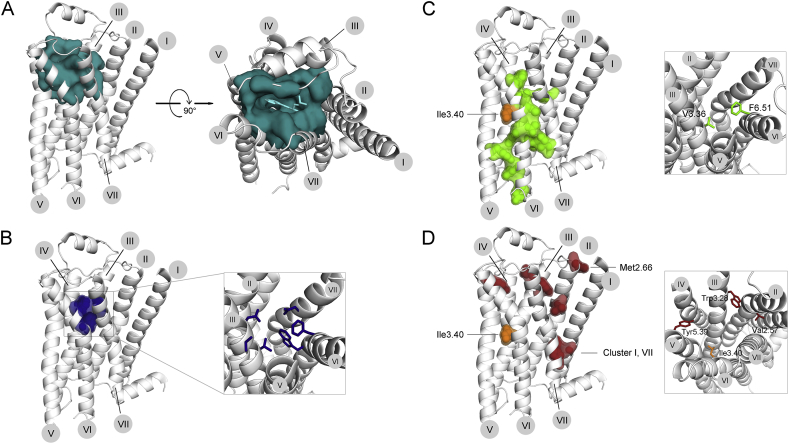
References
-
- Fredriksson R., Lagerstrom M.C., Lundin L.G., Schioth H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol Pharmacol. 2003;63:1256–1272. - PubMed
-
- Pugh E.N., Jr., Lamb T.D. Phototransduction in vertebrate rods and cones: molecular mechanisms of amplification, recovery and light adaptation. In: Stavenga D.G., DeGrip W.J., Pugh E.N. Jr., editors. Handbook of Biological Physics. Molecular Mechanisms in Visual Transduction. Elsevier; Amsterdam: 2000. pp. 183–255.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources