

Phospho1 deficiency transiently modifies bone architecture yet produces consistent modification in osteocyte differentiation and vascular porosity with ageing
- PMID: 26232374
- PMCID: PMC4652607
- DOI: 10.1016/j.bone.2015.07.035
Phospho1 deficiency transiently modifies bone architecture yet produces consistent modification in osteocyte differentiation and vascular porosity with ageing
Abstract
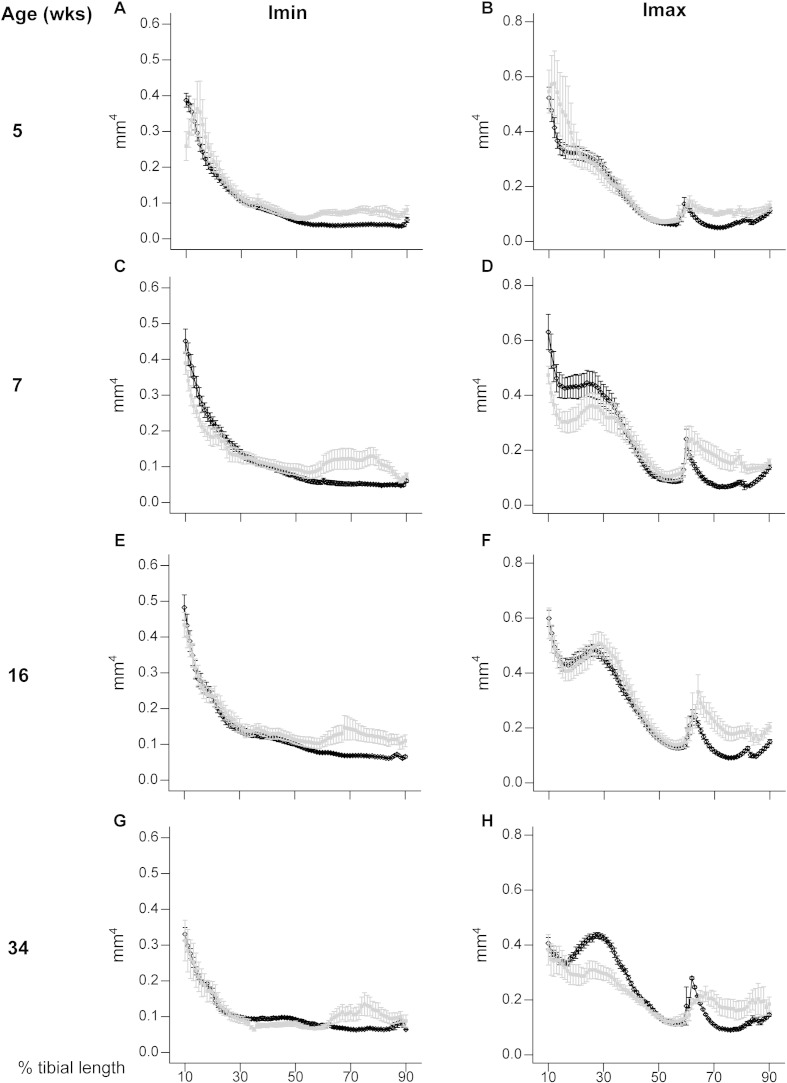
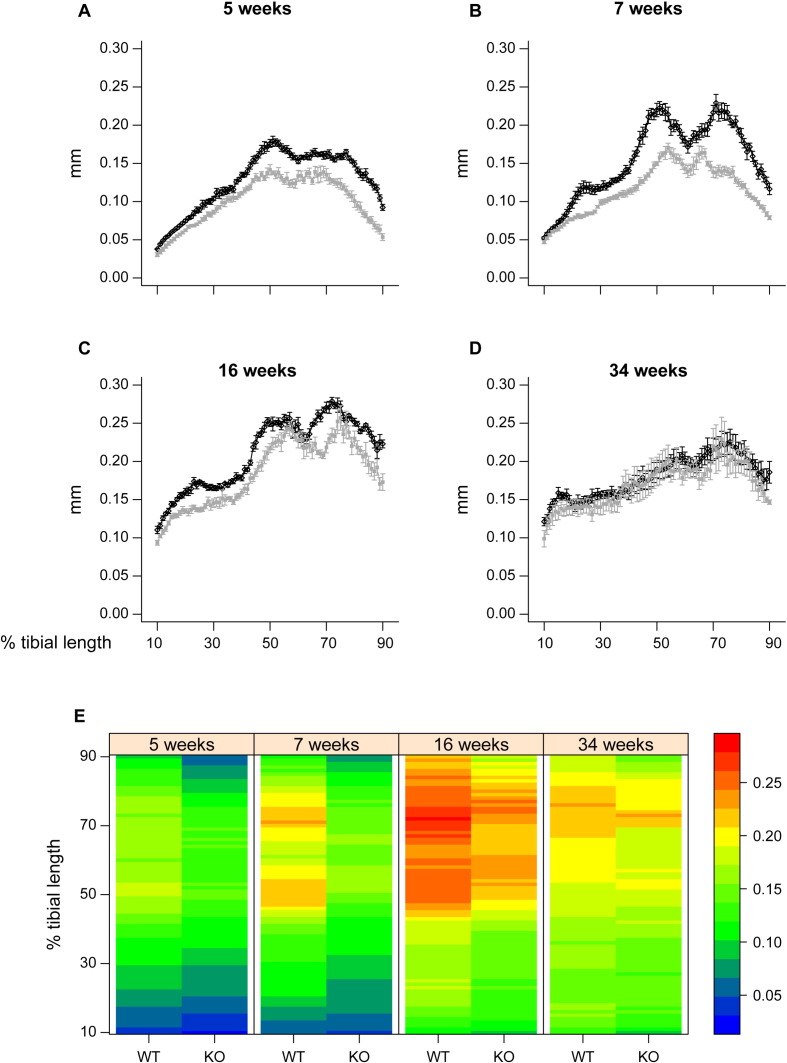
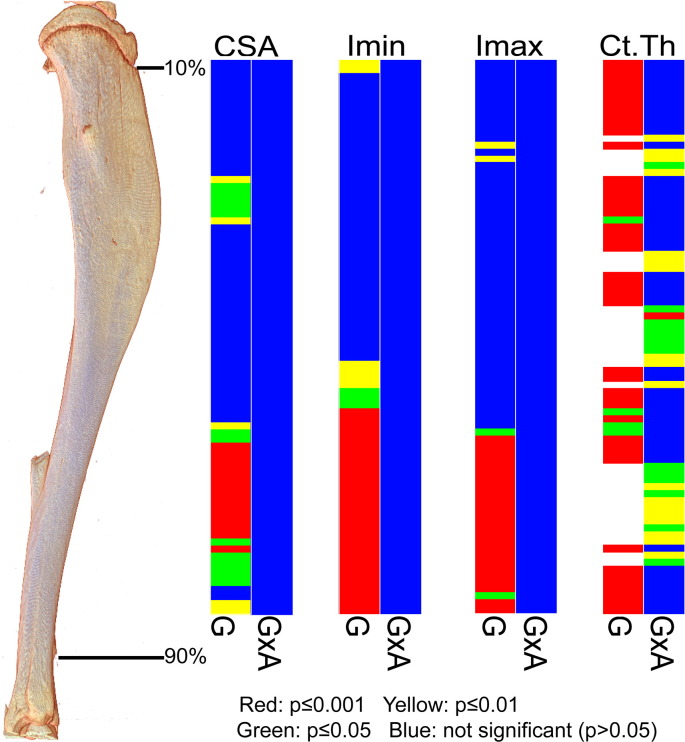
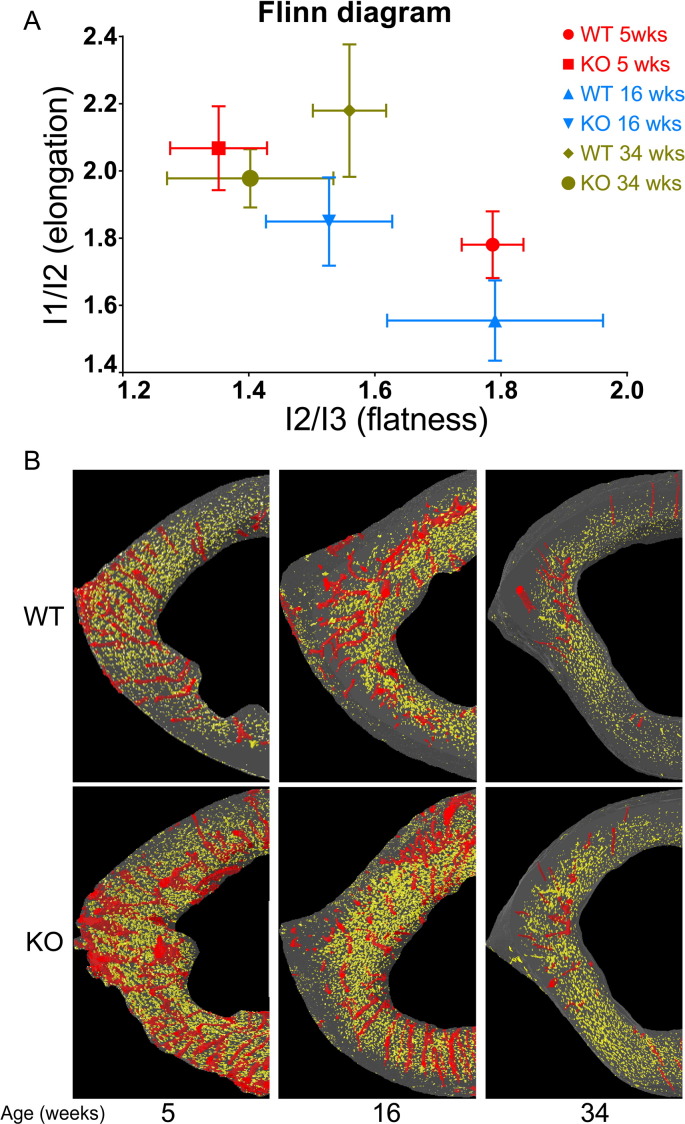
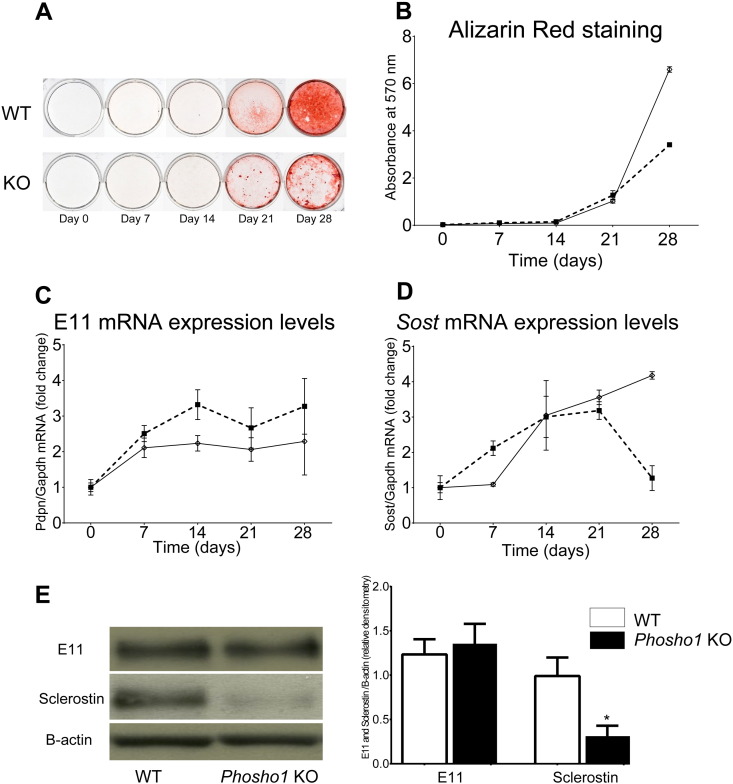
PHOSPHO1 is one of principal proteins involved in initiating bone matrix mineralisation. Recent studies have found that Phospho1 KO mice (Phospho1-R74X) display multiple skeletal abnormalities with spontaneous fractures, bowed long bones, osteomalacia and scoliosis. These analyses have however been limited to young mice and it remains unclear whether the role of PHOSPHO1 is conserved in the mature murine skeleton where bone turnover is limited. In this study, we have used ex-vivo computerised tomography to examine the effect of Phospho1 deletion on tibial bone architecture in mice at a range of ages (5, 7, 16 and 34 weeks of age) to establish whether its role is conserved during skeletal growth and maturation. Matrix mineralisation has also been reported to influence terminal osteoblast differentiation into osteocytes and we have also explored whether hypomineralised bones in Phospho1 KO mice exhibit modified osteocyte lacunar and vascular porosity. Our data reveal that Phospho1 deficiency generates age-related defects in trabecular architecture and compromised cortical microarchitecture with greater porosity accompanied by marked alterations in osteocyte shape, significant increases in osteocytic lacuna and vessel number. Our in vitro studies examining the behaviour of osteoblast derived from Phospho1 KO and wild-type mice reveal reduced levels of matrix mineralisation and modified osteocytogenic programming in cells deficient in PHOSPHO1. Together our data suggest that deficiency in PHOSPHO1 exerts modifications in bone architecture that are transient and depend upon age, yet produces consistent modification in lacunar and vascular porosity. It is possible that the inhibitory role of PHOSPHO1 on osteocyte differentiation leads to these age-related changes in bone architecture. It is also intriguing to note that this apparent acceleration in osteocyte differentiation evident in the hypomineralised bones of Phospho1 KO mice suggests an uncoupling of the interplay between osteocytogenesis and biomineralisation. Further studies are required to dissect the molecular processes underlying the regulatory influences exerted by PHOSPHO1 on the skeleton with ageing.
Keywords: MicroCT; Mineralisation; Osteoblast; Osteocyte; PHOSPHO1; Vascular porosity.
Copyright © 2015. Published by Elsevier Inc.
Figures
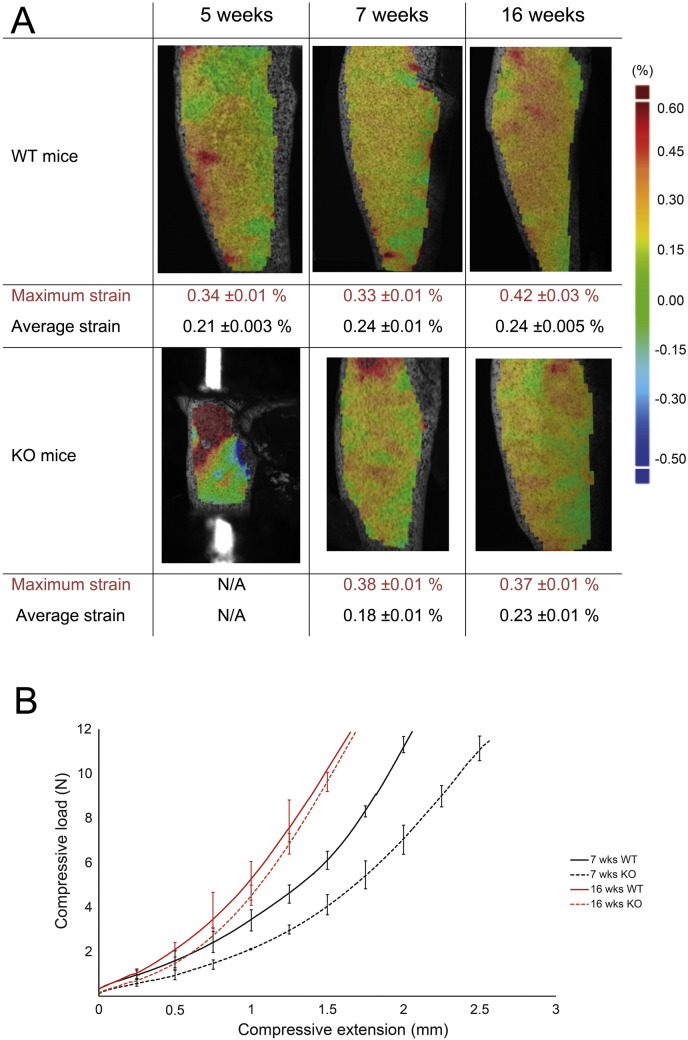
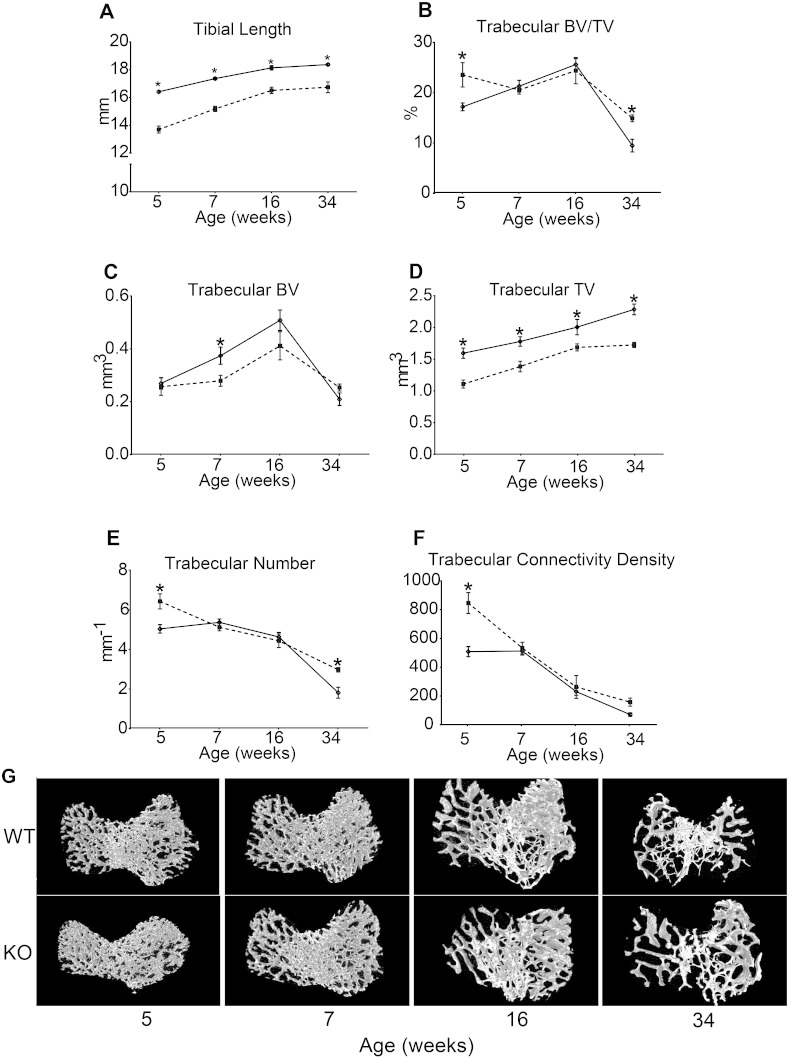
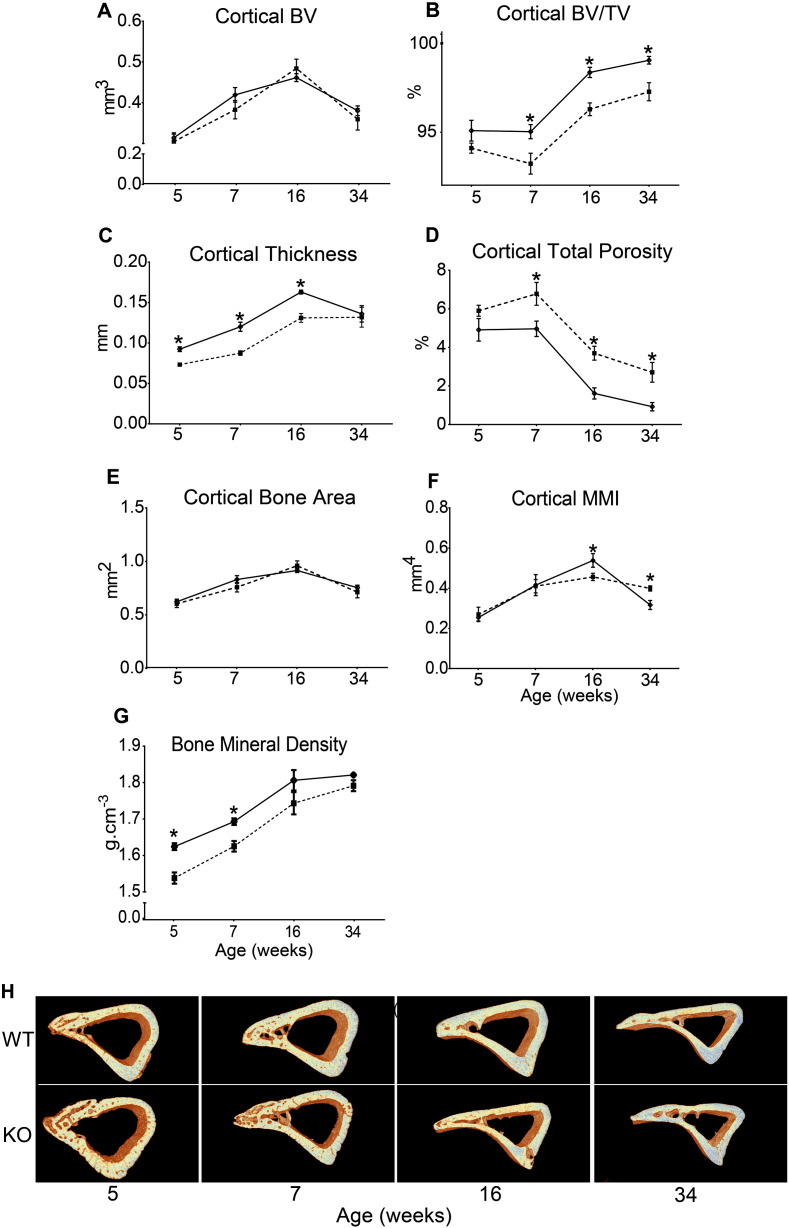
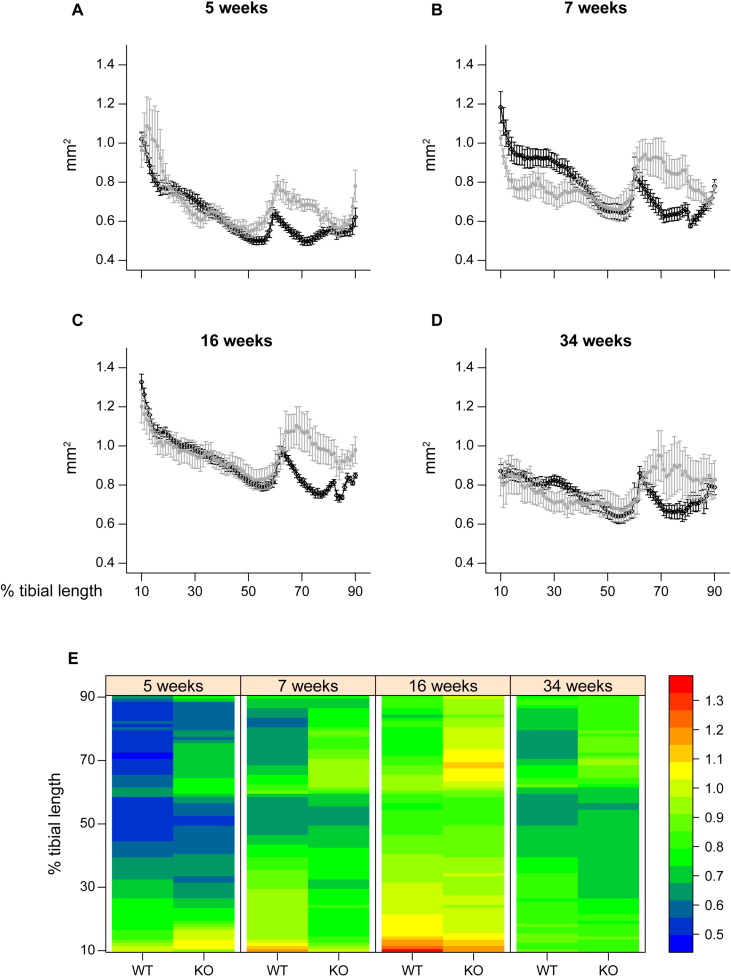
References
-
- Kawasaki K., Buchanan A.V., Weiss K.M. Biomineralization in humans: making the hard choices in life. Annu. Rev. Genet. 2009;43:119–142. - PubMed
-
- Anderson H.C., Garimella R., Tague S.E. The role of matrix vesicles in growth plate development and biomineralization. Front. Biosci. 2005;10:822–837. - PubMed
-
- Boskey A.L., Maresca M., Ullrich W., Doty S.B., Butler W.T., Prince C.W. Osteopontin–hydroxyapatite interactions in vitro: inhibition of hydroxyapatite formation and growth in a gelatin-gel. Bone Miner. 1993;22:147–159. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
