

Age-related changes in dentate gyrus cell numbers, neurogenesis, and associations with cognitive impairments in the rhesus monkey
- PMID: 26236203
- PMCID: PMC4500920
- DOI: 10.3389/fnsys.2015.00102
Age-related changes in dentate gyrus cell numbers, neurogenesis, and associations with cognitive impairments in the rhesus monkey
Abstract
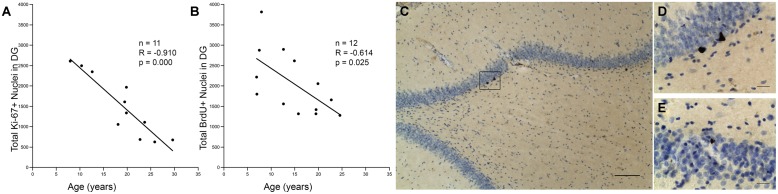
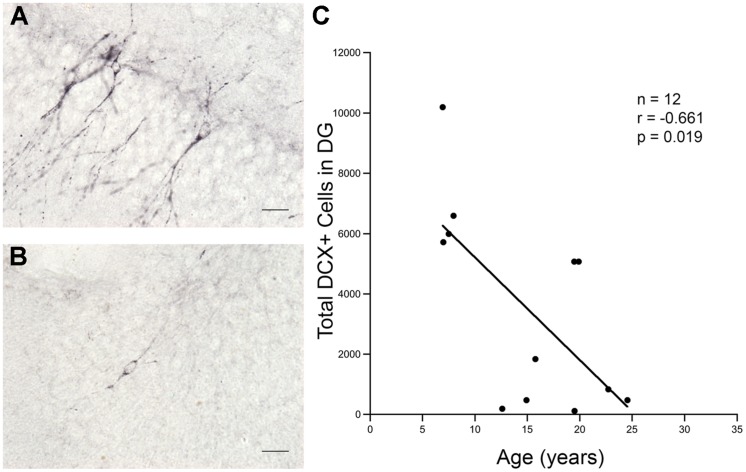
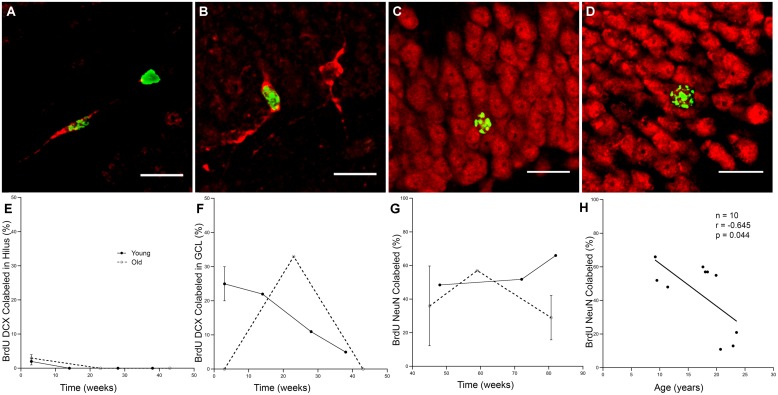
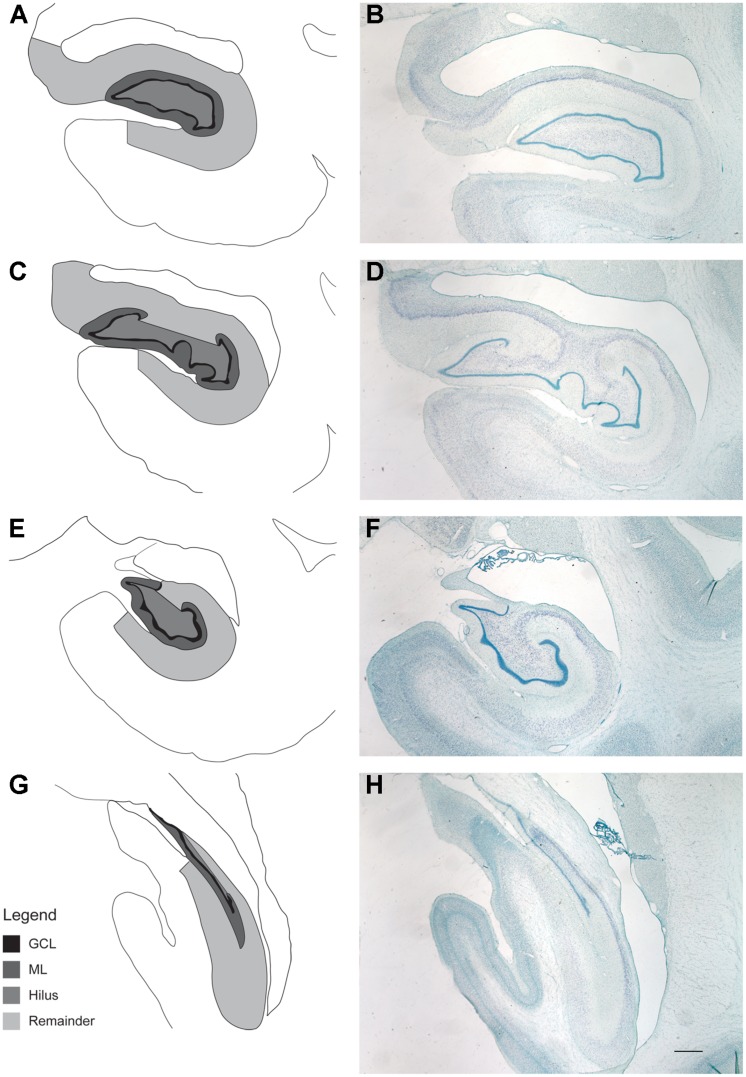
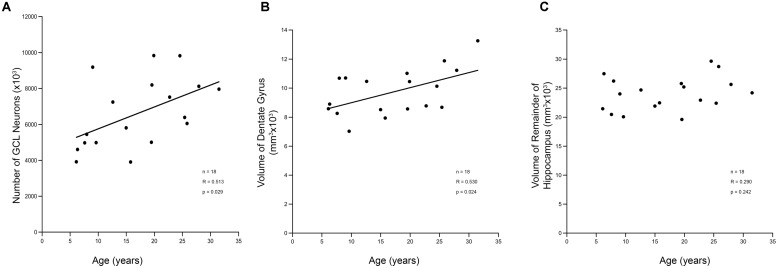
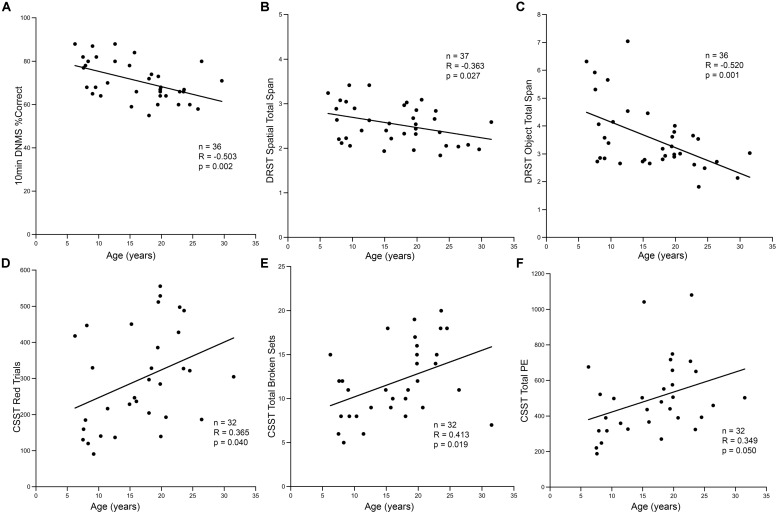
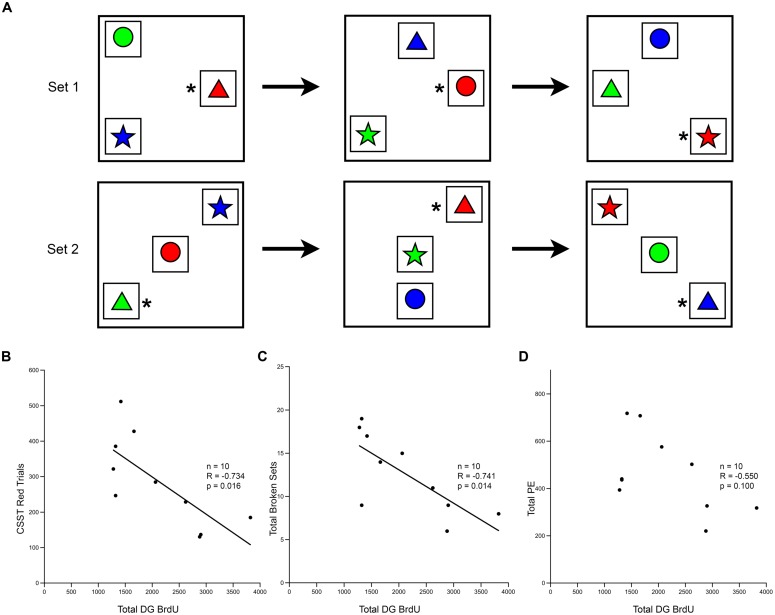
The generation of new neurons in the adult mammalian brain is well-established for the hippocampal dentate gyrus (DG). However, the role of neurogenesis in hippocampal function and cognition, how it changes in aging, and the mechanisms underlying this are yet to be elucidated in the monkey brain. To address this, we investigated adult neurogenesis in the DG of 42 rhesus monkeys (39 cognitively tested) ranging in age from young adult to the elderly. We report here that there is an age-related decline in proliferation and a delayed development of adult neuronal phenotype. Additionally, we show that many of the new neurons survive throughout the lifetime of the animal and may contribute to a modest increase in total neuron number in the granule cell layer of the DG over the adult life span. Lastly, we find that measures of decreased adult neurogenesis are only modestly predictive of age-related cognitive impairment.
Keywords: adult neurogenesis; bromodeoxyuridine; cognitive aging; dentate gyrus; doublecortin; learning; non-human primate; stereology.
Figures
Similar articles
-
Quantitative analysis of postnatal neurogenesis and neuron number in the macaque monkey dentate gyrus.Eur J Neurosci. 2010 Jan;31(2):273-85. doi: 10.1111/j.1460-9568.2009.07061.x. Epub 2010 Jan 13. Eur J Neurosci. 2010. PMID: 20074220 Free PMC article.
-
Age-Dependent Remarkable Regenerative Potential of the Dentate Gyrus Provided by Intrinsic Stem Cells.J Neurosci. 2020 Jan 29;40(5):974-995. doi: 10.1523/JNEUROSCI.1010-19.2019. Epub 2020 Jan 20. J Neurosci. 2020. PMID: 31959697 Free PMC article.
-
Hippocampal granule cell loss in human chronic alcohol abusers.Neurobiol Dis. 2018 Dec;120:63-75. doi: 10.1016/j.nbd.2018.08.011. Epub 2018 Sep 3. Neurobiol Dis. 2018. PMID: 30189262
-
How to make a hippocampal dentate gyrus granule neuron.Development. 2014 Jun;141(12):2366-75. doi: 10.1242/dev.096776. Development. 2014. PMID: 24917496 Review.
-
Adult neurogenesis in the mammalian dentate gyrus.Anat Histol Embryol. 2020 Jan;49(1):3-16. doi: 10.1111/ahe.12496. Epub 2019 Sep 30. Anat Histol Embryol. 2020. PMID: 31568602 Review.
Cited by
-
Adult neurogenesis in the primate hippocampus.Zool Res. 2023 Mar 18;44(2):315-322. doi: 10.24272/j.issn.2095-8137.2022.399. Zool Res. 2023. PMID: 36785898 Free PMC article. Review.
-
Age-Related Changes in Synaptic Plasticity Associated with Mossy Fiber Terminal Integration during Adult Neurogenesis.eNeuro. 2020 May 20;7(3):ENEURO.0030-20.2020. doi: 10.1523/ENEURO.0030-20.2020. Print 2020 May/Jun. eNeuro. 2020. PMID: 32332082 Free PMC article.
-
Evaluation of Long-Term Cryostorage of Brain Tissue Sections for Quantitative Histochemistry.J Histochem Cytochem. 2017 Mar;65(3):153-171. doi: 10.1369/0022155416686934. Epub 2017 Jan 12. J Histochem Cytochem. 2017. PMID: 28080173 Free PMC article.
-
Mesenchymal-derived extracellular vesicles enhance microglia-mediated synapse remodeling after cortical injury in aging Rhesus monkeys.J Neuroinflammation. 2023 Sep 2;20(1):201. doi: 10.1186/s12974-023-02880-0. J Neuroinflammation. 2023. PMID: 37660145 Free PMC article.
-
Human Hippocampal Neurogenesis Persists throughout Aging.Cell Stem Cell. 2018 Apr 5;22(4):589-599.e5. doi: 10.1016/j.stem.2018.03.015. Cell Stem Cell. 2018. PMID: 29625071 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources