

Brassinosteroids Are Master Regulators of Gibberellin Biosynthesis in Arabidopsis
- PMID: 26243314
- PMCID: PMC4568508
- DOI: 10.1105/tpc.15.00433
Brassinosteroids Are Master Regulators of Gibberellin Biosynthesis in Arabidopsis
Abstract
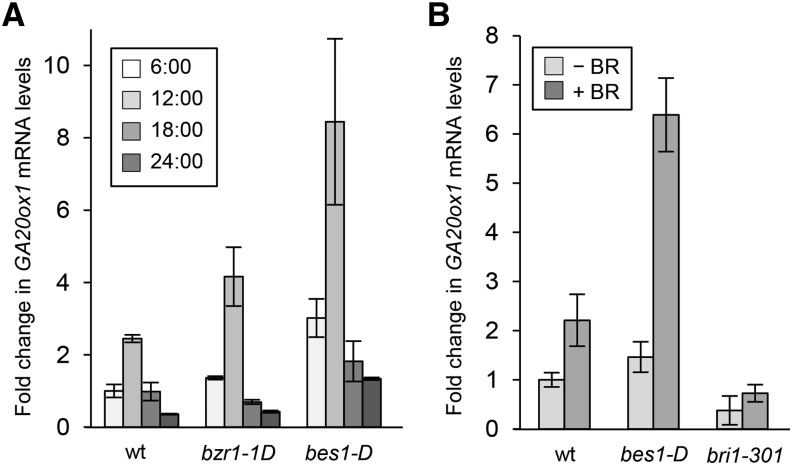
Plant growth and development are highly regulated processes that are coordinated by hormones including the brassinosteroids (BRs), a group of steroids with structural similarity to steroid hormones of mammals. Although it is well understood how BRs are produced and how their signals are transduced, BR targets, which directly confer the hormone's growth-promoting effects, have remained largely elusive. Here, we show that BRs regulate the biosynthesis of gibberellins (GAs), another class of growth-promoting hormones, in Arabidopsis thaliana. We reveal that Arabidopsis mutants deficient in BR signaling are severely impaired in the production of bioactive GA, which is correlated with defective GA biosynthetic gene expression. Expression of the key GA biosynthesis gene GA20ox1 in the BR signaling mutant bri1-301 rescues many of its developmental defects. We provide evidence that supports a model in which the BR-regulated transcription factor BES1 binds to a regulatory element in promoters of GA biosynthesis genes in a BR-induced manner to control their expression. In summary, our study underscores a role of BRs as master regulators of GA biosynthesis and shows that this function is of major relevance for the growth and development of vascular plants.
© 2015 American Society of Plant Biologists. All rights reserved.
Figures







Comment in
-
Taking Hormone Crosstalk to a New Level: Brassinosteroids Regulate Gibberellin Biosynthesis.Plant Cell. 2015 Aug;27(8):2081. doi: 10.1105/tpc.15.00700. Epub 2015 Aug 14. Plant Cell. 2015. PMID: 26276835 Free PMC article. No abstract available.
-
Interactions between Brassinosteroids and Gibberellins: Synthesis or Signaling?Plant Cell. 2016 Apr;28(4):829-32. doi: 10.1105/tpc.15.00917. Epub 2016 Mar 22. Plant Cell. 2016. PMID: 27006485 Free PMC article. No abstract available.
-
Reply: Interaction between Brassinosteroids and Gibberellins: Synthesis or Signaling? In Arabidopsis, Both!Plant Cell. 2016 Apr;28(4):836-9. doi: 10.1105/tpc.16.00120. Epub 2016 Mar 22. Plant Cell. 2016. PMID: 27006486 Free PMC article. No abstract available.
Similar articles
-
An interaction between BZR1 and DELLAs mediates direct signaling crosstalk between brassinosteroids and gibberellins in Arabidopsis.Sci Signal. 2012 Oct 2;5(244):ra72. doi: 10.1126/scisignal.2002908. Sci Signal. 2012. PMID: 23033541
-
Transcription factor HAT1 is phosphorylated by BIN2 kinase and mediates brassinosteroid repressed gene expression in Arabidopsis.Plant J. 2014 Jan;77(1):59-70. doi: 10.1111/tpj.12368. Epub 2013 Dec 3. Plant J. 2014. PMID: 24164091
-
Genetic analyses of interactions among gibberellin, abscisic acid, and brassinosteroids in the control of flowering time in Arabidopsis thaliana.PLoS One. 2010 Nov 17;5(11):e14012. doi: 10.1371/journal.pone.0014012. PLoS One. 2010. PMID: 21103336 Free PMC article.
-
Molecular Mechanisms of Brassinosteroid-Mediated Responses to Changing Environments in Arabidopsis.Int J Mol Sci. 2020 Apr 15;21(8):2737. doi: 10.3390/ijms21082737. Int J Mol Sci. 2020. PMID: 32326491 Free PMC article. Review.
-
Plants grow on brassinosteroids.Curr Opin Plant Biol. 2011 Oct;14(5):530-7. doi: 10.1016/j.pbi.2011.05.004. Epub 2011 Jul 27. Curr Opin Plant Biol. 2011. PMID: 21802346 Review.
Cited by
-
Brassinosteroids, the Sixth Class of Phytohormones: A Molecular View from the Discovery to Hormonal Interactions in Plant Development and Stress Adaptation.Int J Mol Sci. 2019 Jan 15;20(2):331. doi: 10.3390/ijms20020331. Int J Mol Sci. 2019. PMID: 30650539 Free PMC article. Review.
-
Transcription factor BES1 interacts with HSFA1 to promote heat stress resistance of plants.EMBO J. 2022 Feb 1;41(3):e108664. doi: 10.15252/embj.2021108664. Epub 2022 Jan 4. EMBO J. 2022. PMID: 34981847 Free PMC article.
-
Regulatory Effects of ABA and GA on the Expression of Conglutin Genes and LAFL Network Genes in Yellow Lupine (Lupinus luteus L.) Seeds.Int J Mol Sci. 2023 Aug 3;24(15):12380. doi: 10.3390/ijms241512380. Int J Mol Sci. 2023. PMID: 37569754 Free PMC article.
-
Hormonal control of cold stress responses in plants.Cell Mol Life Sci. 2016 Feb;73(4):797-810. doi: 10.1007/s00018-015-2089-6. Epub 2015 Nov 23. Cell Mol Life Sci. 2016. PMID: 26598281 Free PMC article. Review.
-
Barley Brassinosteroid Mutants Provide an Insight into Phytohormonal Homeostasis in Plant Reaction to Drought Stress.Front Plant Sci. 2016 Dec 2;7:1824. doi: 10.3389/fpls.2016.01824. eCollection 2016. Front Plant Sci. 2016. PMID: 27994612 Free PMC article.
References
-
- Alboresi A., Gestin C., Leydecker M.T., Bedu M., Meyer C., Truong H.N. (2005). Nitrate, a signal relieving seed dormancy in Arabidopsis. Plant Cell Environ. 28: 500–512. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
