

Identification and Mechanistic Analysis of a Novel Tick-Derived Inhibitor of Thrombin
- PMID: 26244557
- PMCID: PMC4526366
- DOI: 10.1371/journal.pone.0133991
Identification and Mechanistic Analysis of a Novel Tick-Derived Inhibitor of Thrombin
Abstract
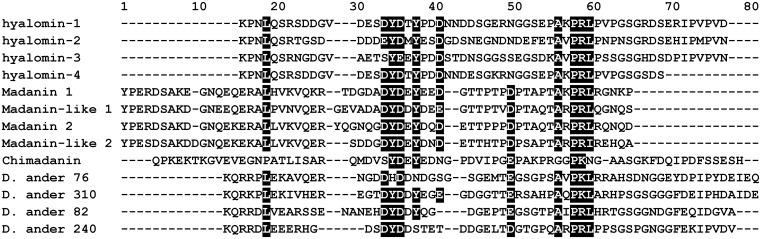
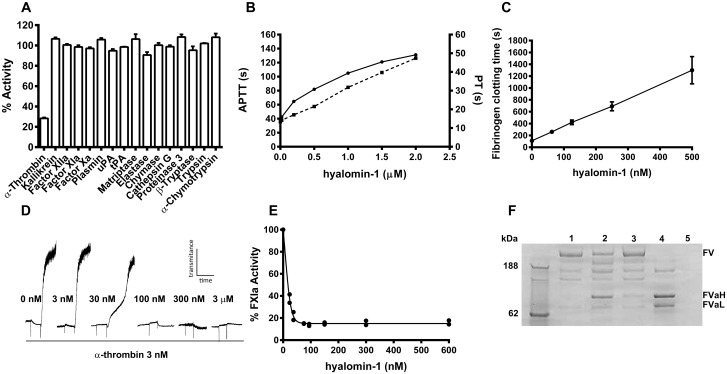
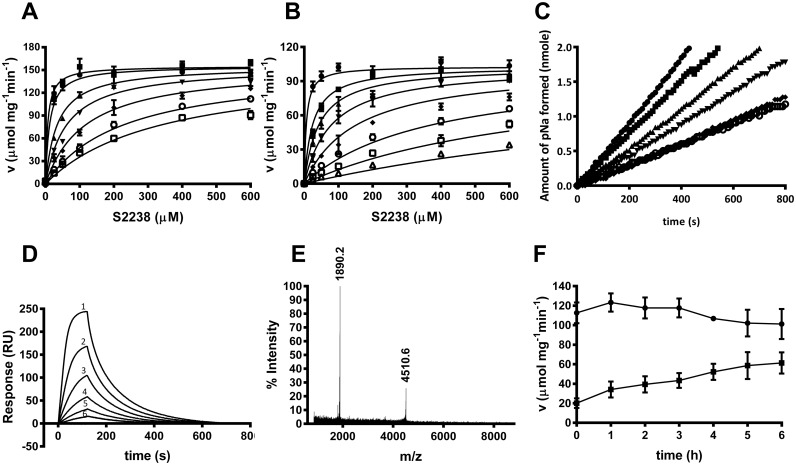
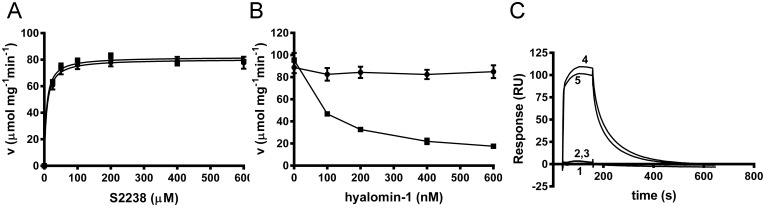
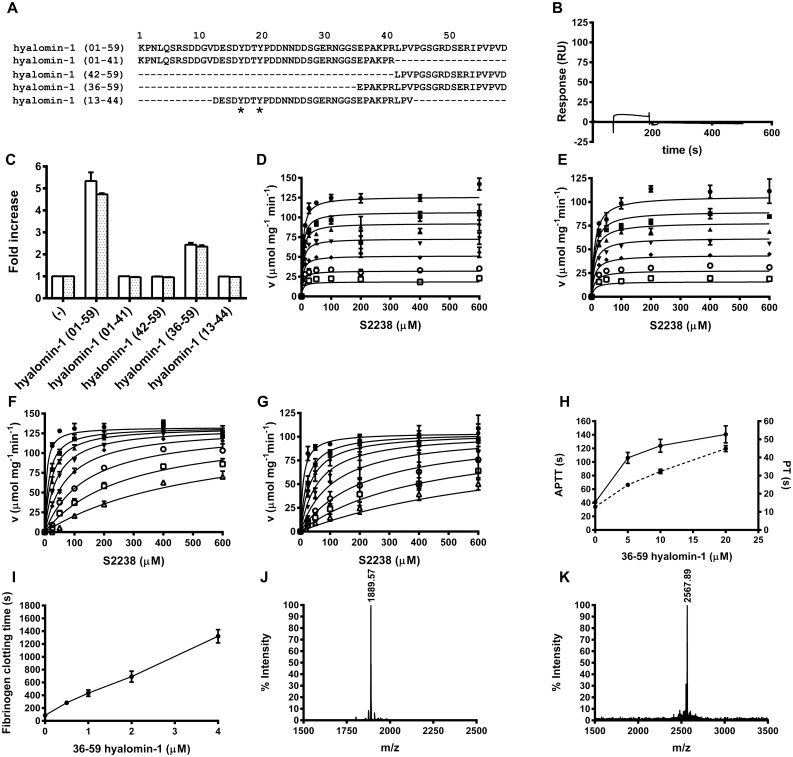
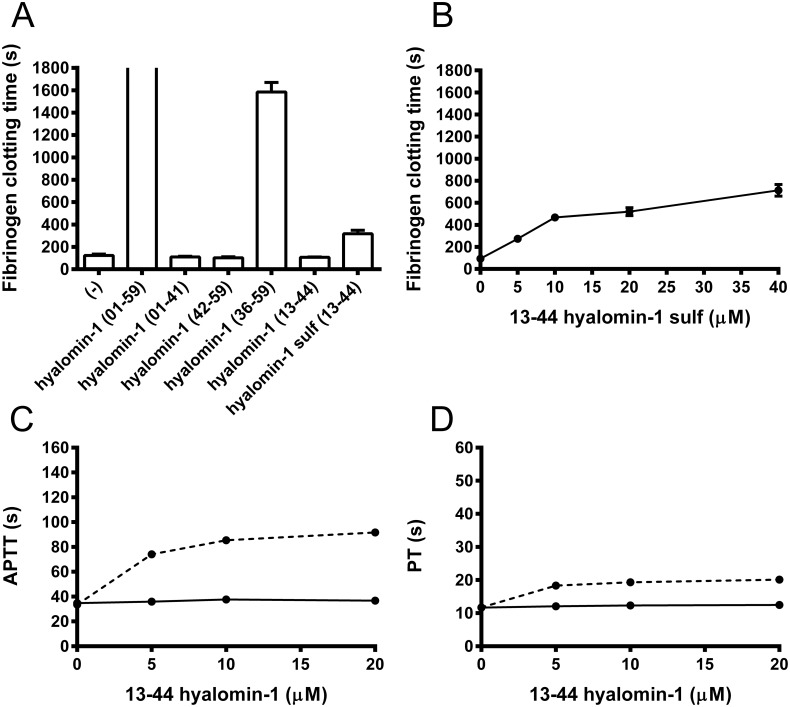
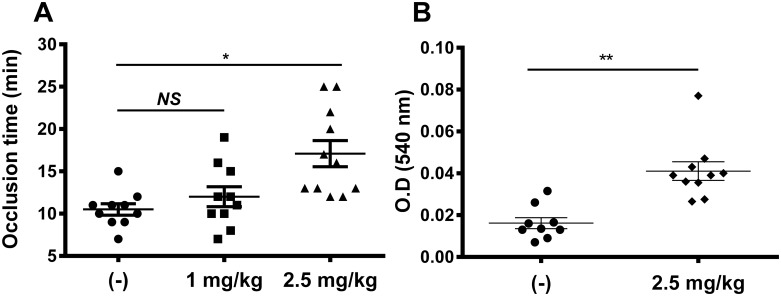
A group of peptides from the salivary gland of the tick Hyalomma marginatum rufipes, a vector of Crimean Congo hemorrhagic fever show weak similarity to the madanins, a group of thrombin-inhibitory peptides from a second tick species, Haemaphysalis longicornis. We have evaluated the anti-serine protease activity of one of these H. marginatum peptides that has been given the name hyalomin-1. Hyalomin-1 was found to be a selective inhibitor of thrombin, blocking coagulation of plasma and inhibiting S2238 hydrolysis in a competitive manner with an inhibition constant (Ki) of 12 nM at an ionic strength of 150 mM. It also blocks the thrombin-mediated activation of coagulation factor XI, thrombin-mediated platelet aggregation, and the activation of coagulation factor V by thrombin. Hyalomin-1 is cleaved at a canonical thrombin cleavage site but the cleaved products do not inhibit coagulation. However, the C-terminal cleavage product showed non-competitive inhibition of S2238 hydrolysis. A peptide combining the N-terminal parts of the molecule with the cleavage region did not interact strongly with thrombin, but a 24-residue fragment containing the cleavage region and the C-terminal fragment inhibited the enzyme in a competitive manner and also inhibited coagulation of plasma. These results suggest that the peptide acts by binding to the active site as well as exosite I or the autolysis loop of thrombin. Injection of 2.5 mg/kg of hyalomin-1 increased arterial occlusion time in a mouse model of thrombosis, suggesting this peptide could be a candidate for clinical use as an antithrombotic.
Conflict of interest statement
Figures
Similar articles
-
Identification and characterization of novel salivary thrombin inhibitors from the ixodidae tick, Haemaphysalis longicornis.Eur J Biochem. 2003 May;270(9):1926-34. doi: 10.1046/j.1432-1033.2003.03560.x. Eur J Biochem. 2003. PMID: 12709051
-
Expression and characterization of haemathrins, madanin-like thrombin inhibitors, isolated from the salivary gland of tick Haemaphysalis bispinosa (Acari: Ixodidae).Thromb Res. 2017 Apr;152:20-29. doi: 10.1016/j.thromres.2017.01.012. Epub 2017 Jan 31. Thromb Res. 2017. PMID: 28213103
-
Inhibitors of serine proteinases from blood coagulation cascade - view on current developments.Mini Rev Med Chem. 2012 Jul;12(8):721-30. doi: 10.2174/138955712801264819. Mini Rev Med Chem. 2012. PMID: 22512557 Review.
-
Avathrin: a novel thrombin inhibitor derived from a multicopy precursor in the salivary glands of the ixodid tick, Amblyomma variegatum.FASEB J. 2017 Jul;31(7):2981-2995. doi: 10.1096/fj.201601216R. Epub 2017 Mar 31. FASEB J. 2017. PMID: 28363953
-
[Peptides as inhibitors of thrombin coagulation activity and of thrombocyte aggregation].Usp Fiziol Nauk. 1999 Apr-Jun;30(2):80-91. Usp Fiziol Nauk. 1999. PMID: 10420478 Review. Russian.
Cited by
-
Host Immune Responses to Salivary Components - A Critical Facet of Tick-Host Interactions.Front Cell Infect Microbiol. 2022 Mar 16;12:809052. doi: 10.3389/fcimb.2022.809052. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35372098 Free PMC article. Review.
-
An Enabling Peptide Ligation Induced by Thiol-Salicylaldehyde Ester for Chemical Protein Synthesis.Adv Sci (Weinh). 2024 Dec;11(46):e2408538. doi: 10.1002/advs.202408538. Epub 2024 Oct 23. Adv Sci (Weinh). 2024. PMID: 39440515 Free PMC article.
-
Interactions of Human Dermal Dendritic Cells and Langerhans Cells Treated with Hyalomma Tick Saliva with Crimean-Congo Hemorrhagic Fever Virus.Viruses. 2018 Jul 20;10(7):381. doi: 10.3390/v10070381. Viruses. 2018. PMID: 30036960 Free PMC article.
-
Rapid assembly and profiling of an anticoagulant sulfoprotein library.Proc Natl Acad Sci U S A. 2019 Jul 9;116(28):13873-13878. doi: 10.1073/pnas.1905177116. Epub 2019 Jun 20. Proc Natl Acad Sci U S A. 2019. PMID: 31221752 Free PMC article.
-
Purification and characterization of thrombin from camel plasma: interaction with camel tick salivary gland thrombin inhibitor.J Genet Eng Biotechnol. 2023 Jan 23;21(1):7. doi: 10.1186/s43141-023-00464-2. J Genet Eng Biotechnol. 2023. PMID: 36689046 Free PMC article.
References
-
- Bode W (2006) The structure of thrombin: a janus-headed proteinase. Semin Thromb Hemost 32 Suppl 1: 16–31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
