

Enhanced B-Cell Receptor Recognition of the Autoantigen Transglutaminase 2 by Efficient Catalytic Self-Multimerization
- PMID: 26244572
- PMCID: PMC4526674
- DOI: 10.1371/journal.pone.0134922
Enhanced B-Cell Receptor Recognition of the Autoantigen Transglutaminase 2 by Efficient Catalytic Self-Multimerization
Abstract
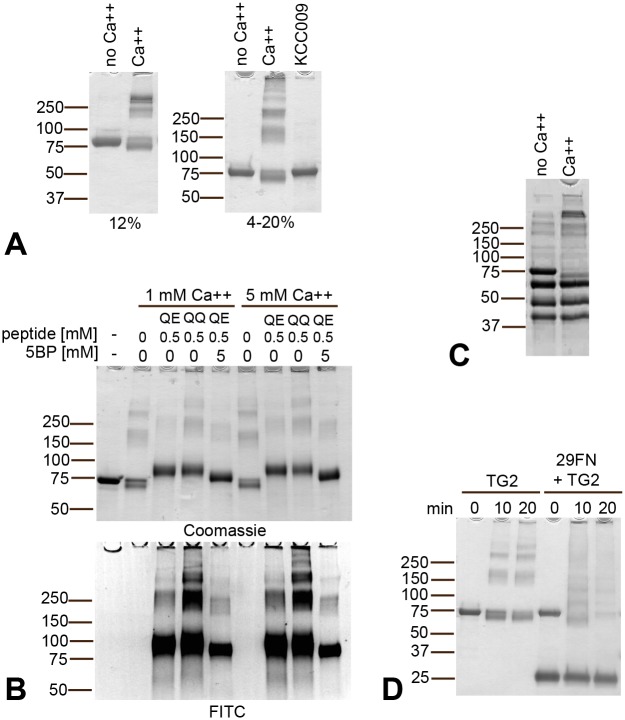
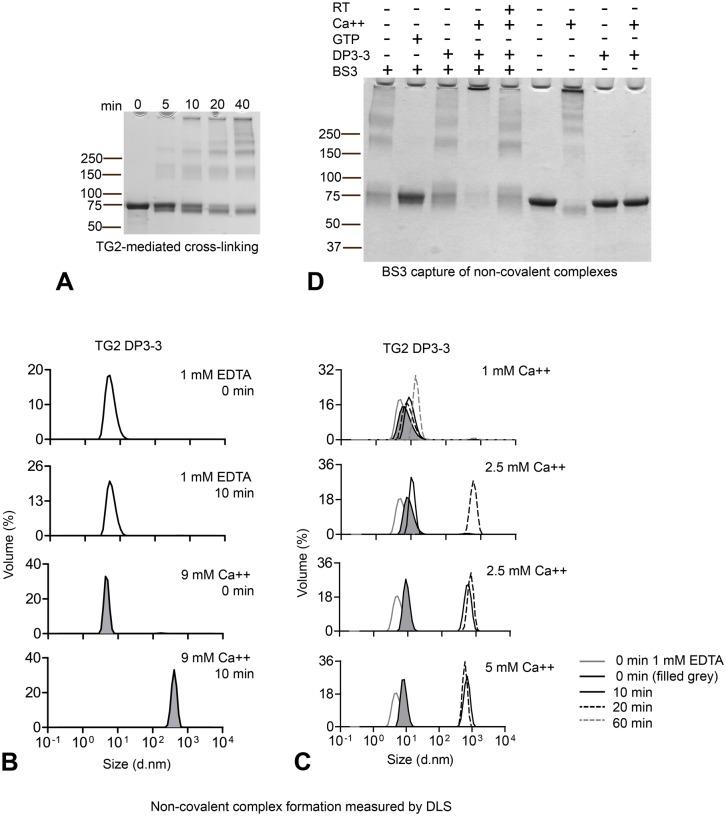
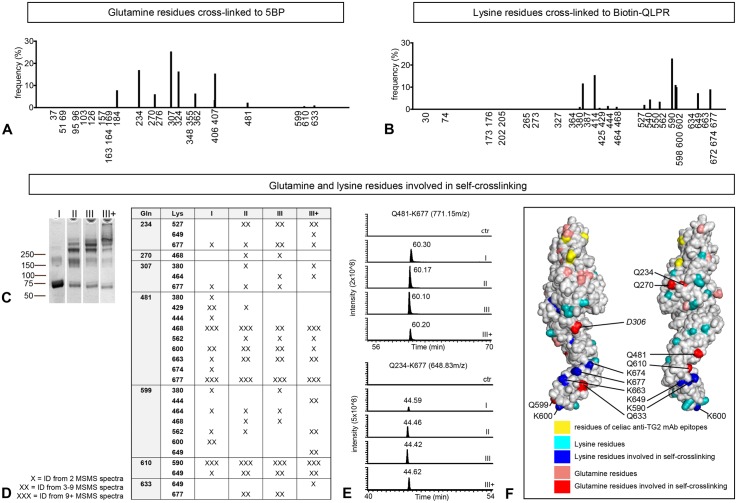
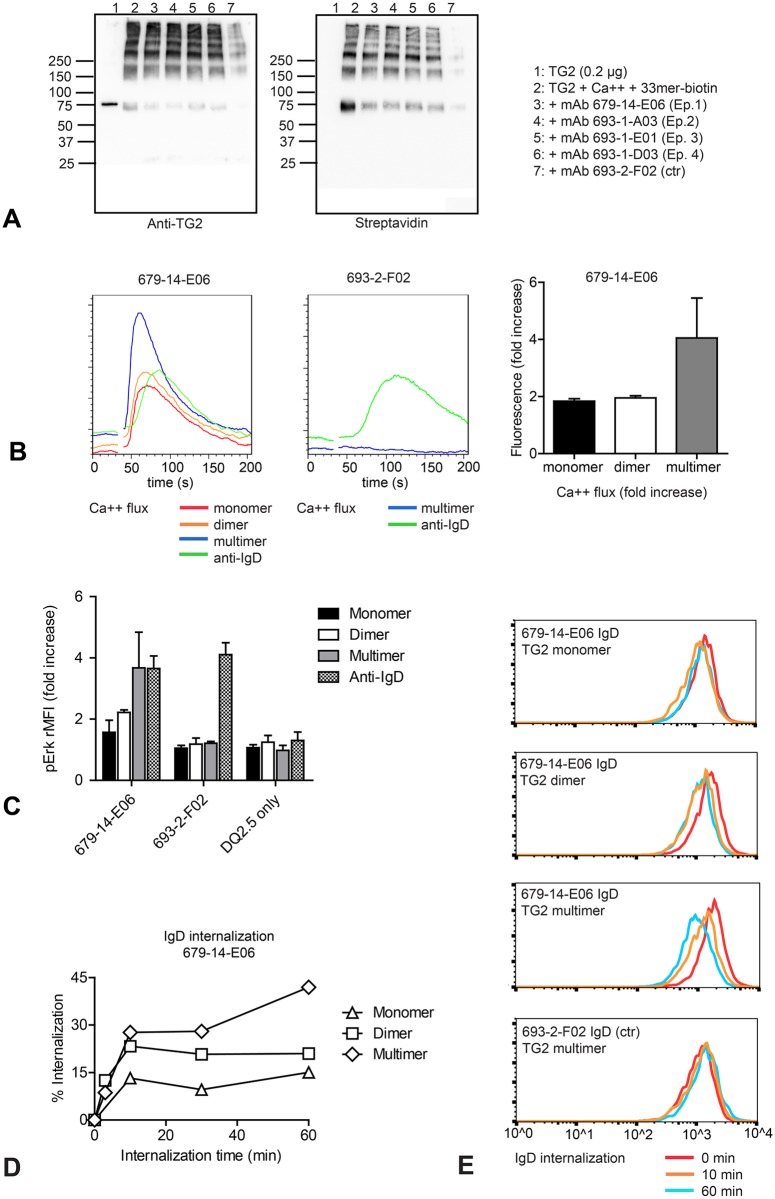
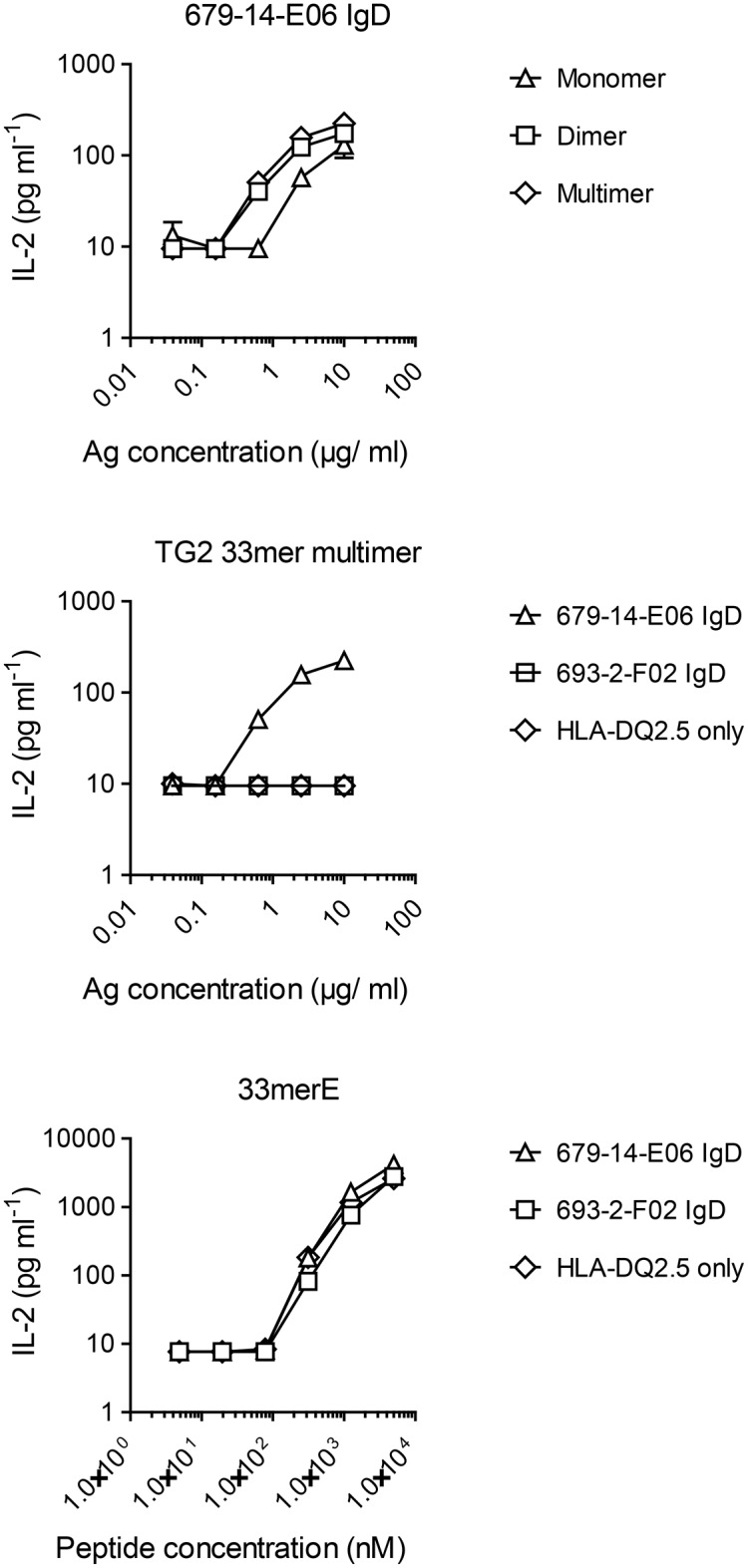
A hallmark of the gluten-driven enteropathy celiac disease is autoantibody production towards the enzyme transglutaminase 2 (TG2) that catalyzes the formation of covalent protein-protein cross-links. Activation of TG2-specific B cells likely involves gluten-specific CD4 T cells as production of the antibodies is dependent on disease-associated HLA-DQ allotypes and dietary intake of gluten. IgA plasma cells producing TG2 antibodies with few mutations are abundant in the celiac gut lesion. These plasma cells and serum antibodies to TG2 drop rapidly after initiation of a gluten-free diet, suggestive of extrafollicular responses or germinal center reactions of short duration. High antigen avidity is known to promote such responses, and is also important for breakage of self-tolerance. We here inquired whether TG2 avidity could be a feature relevant to celiac disease. Using recombinant enzyme we show by dynamic light scattering and gel electrophoresis that TG2 efficiently utilizes itself as a substrate due to conformation-dependent homotypic association, which involves the C-terminal domains of the enzyme. This leads to the formation of covalently linked TG2 multimers. The presence of exogenous substrate such as gluten peptide does not inhibit TG2 self-cross-linking, but rather results in formation of TG2-TG2-gluten complexes. The celiac disease autoantibody epitopes, clustered in the N-terminal part of TG2, are conserved in the TG2-multimers as determined by mass spectrometry and immunoprecipitation analysis. TG2 multimers are superior to TG2 monomer in activating A20 B cells transduced with TG2-specific B-cell receptor, and uptake of TG2-TG2-gluten multimers leads to efficient activation of gluten-specific T cells. Efficient catalytic self-multimerization of TG2 and generation of multivalent TG2 antigen decorated with gluten peptides suggest a mechanism by which self-reactive B cells are activated to give abundant numbers of plasma cells in celiac disease. Importantly, high avidity of the antigen could explain why TG2-specific plasma cells show signs of an extrafollicular generation pathway.
Conflict of interest statement
Figures

References
-
- Lorand L, Graham RM. Transglutaminases: crosslinking enzymes with pleiotropic functions. Nat Rev Mol Cell Biol. 2003;4(2):140–56. - PubMed
-
- Fleckenstein B, Molberg O, Qiao SW, Schmid DG, von der Mulbe F, Elgstoen K, et al. Gliadin T cell epitope selection by tissue transglutaminase in celiac disease. Role of enzyme specificity and pH influence on the transamidation versus deamidation process. J Biol Chem. 2002;277(37):34109–16. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
