

Rod- and cone-driven responses in mice expressing human L-cone pigment
- PMID: 26245314
- PMCID: PMC4600960
- DOI: 10.1152/jn.00188.2015
Rod- and cone-driven responses in mice expressing human L-cone pigment
Abstract
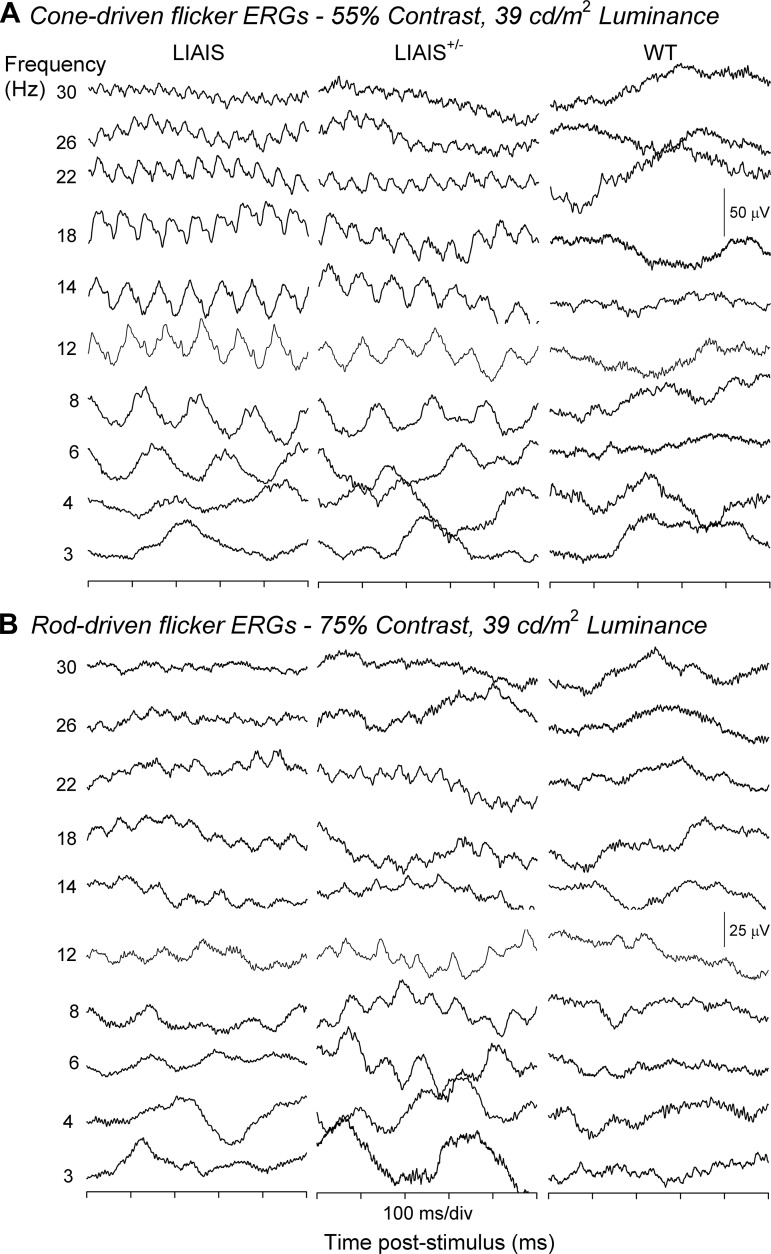
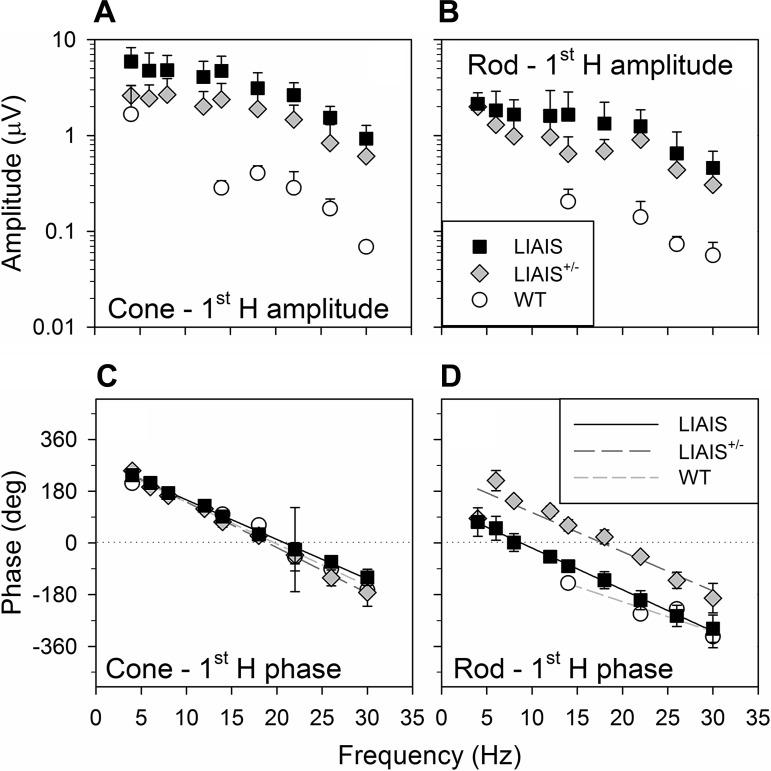
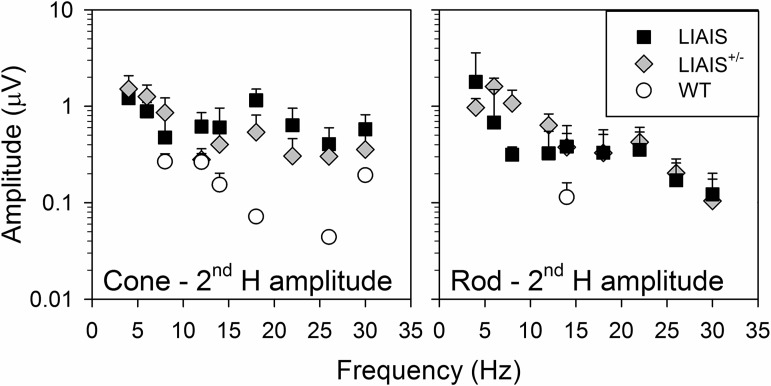
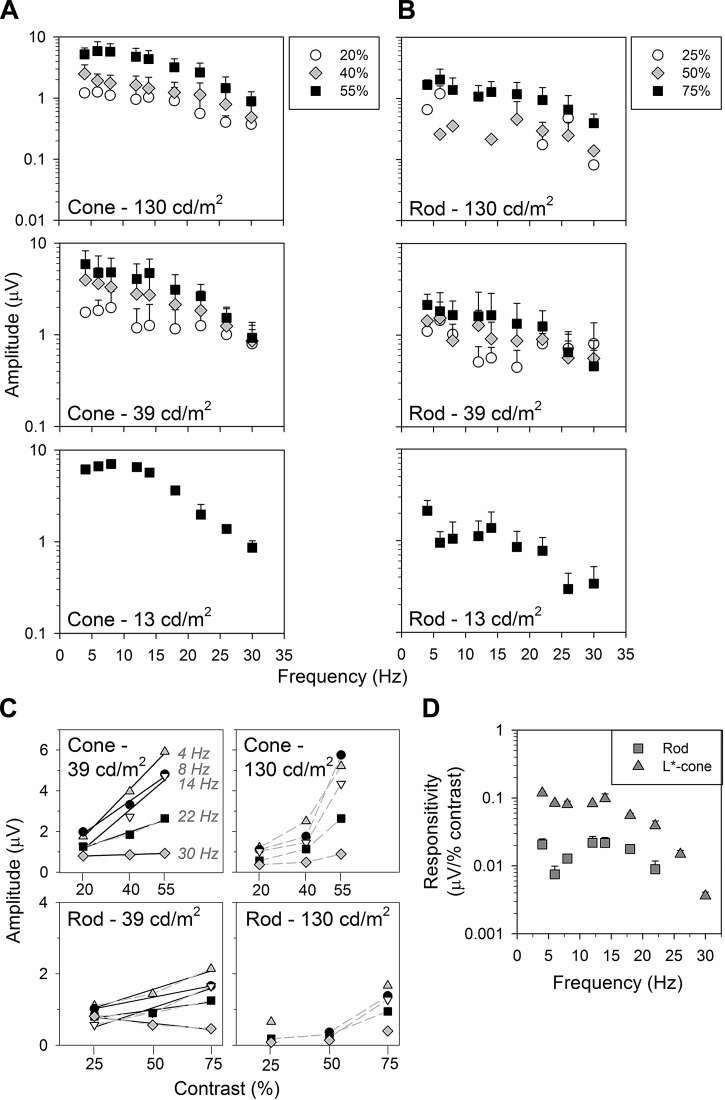
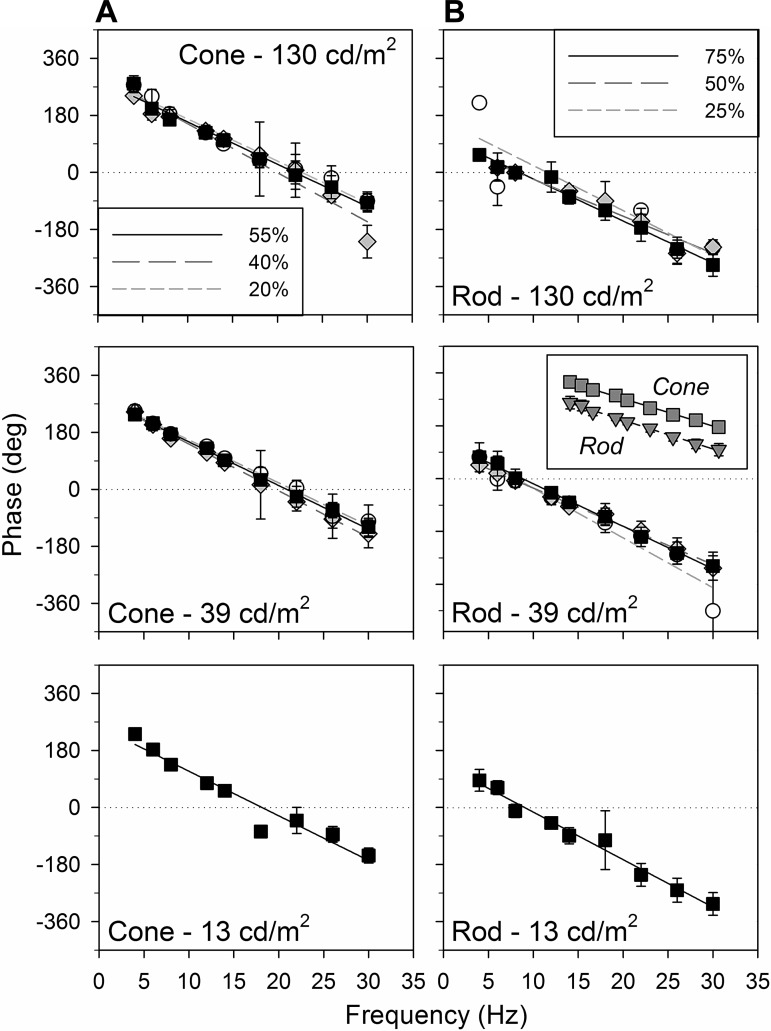
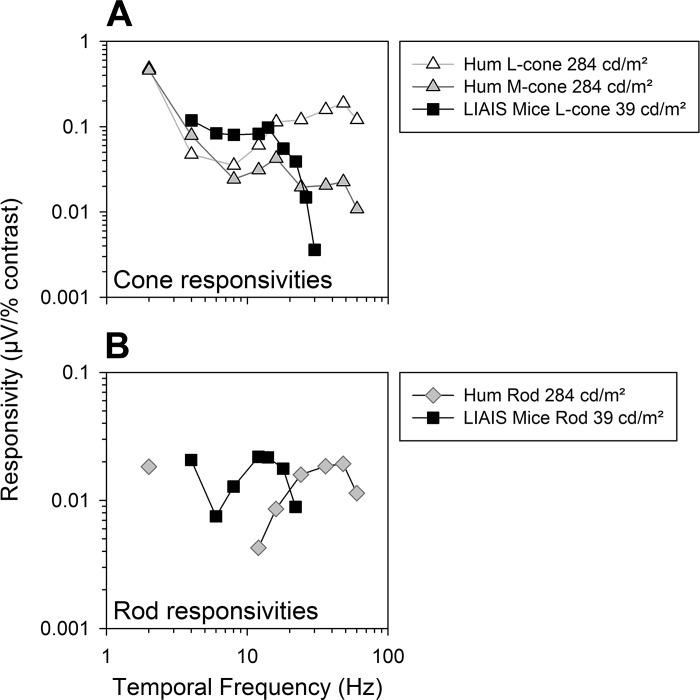
The mouse is commonly used for studying retinal processing, primarily because it is amenable to genetic manipulation. To accurately study photoreceptor driven signals in the healthy and diseased retina, it is of great importance to isolate the responses of single photoreceptor types. This is not easily achieved in mice because of the strong overlap of rod and M-cone absorption spectra (i.e., maxima at 498 and 508 nm, respectively). With a newly developed mouse model (Opn1lw(LIAIS)) expressing a variant of the human L-cone pigment (561 nm) instead of the mouse M-opsin, the absorption spectra are substantially separated, allowing retinal physiology to be studied using silent substitution stimuli. Unlike conventional chromatic isolation methods, this spectral compensation approach can isolate single photoreceptor subtypes without changing the retinal adaptation. We measured flicker electroretinograms in these mutants under ketamine-xylazine sedation with double silent substitution (silent S-cone and either rod or M/L-cones) and obtained robust responses for both rods and (L-)cones. Small signals were yielded in wild-type mice, whereas heterozygotes exhibited responses that were generally intermediate to both. Fundamental response amplitudes and phase behaviors (as a function of temporal frequency) in all genotypes were largely similar. Surprisingly, isolated (L-)cone and rod response properties in the mutant strain were alike. Thus the LIAIS mouse warrants a more comprehensive in vivo assessment of photoreceptor subtype-specific physiology, because it overcomes the hindrance of overlapping spectral sensitivities present in the normal mouse.
Keywords: keywords electrophysiology; mouse; photoreceptors; silent substitution.
Copyright © 2015 the American Physiological Society.
Figures
References
-
- Atorf J, Scholz M, Garreis F, Lehmann J, Brauer L, Kremers J. Functional protective effects of long-term memantine treatment in the DBA/2J mouse. Doc Ophthalmol 126: 221–232, 2013. - PubMed
-
- Bessler P, Klee S, Kellner U, Haueisen J. Silent substitution stimulation of S-cone pathway and L- and M-cone pathway in glaucoma. Invest Ophthalmol Vis Sci 51: 319–326, 2010. - PubMed
-
- Bloomfield SA, Dacheux RF. Rod vision: pathways and processing in the mammalian retina. Prog Retin Eye Res 20: 351–384, 2001. - PubMed
-
- Brainard DH, Calderone JB, Nugent AK, Jacobs GH. Flicker ERG responses to stimuli parametrically modulated in color space. Invest Ophthalmol Vis Sci 40: 2840–2847, 1999. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources