

Neutrophil microvesicles resolve gout by inhibiting C5a-mediated priming of the inflammasome
- PMID: 26245757
- PMCID: PMC4893114
- DOI: 10.1136/annrheumdis-2015-207338
Neutrophil microvesicles resolve gout by inhibiting C5a-mediated priming of the inflammasome
Abstract
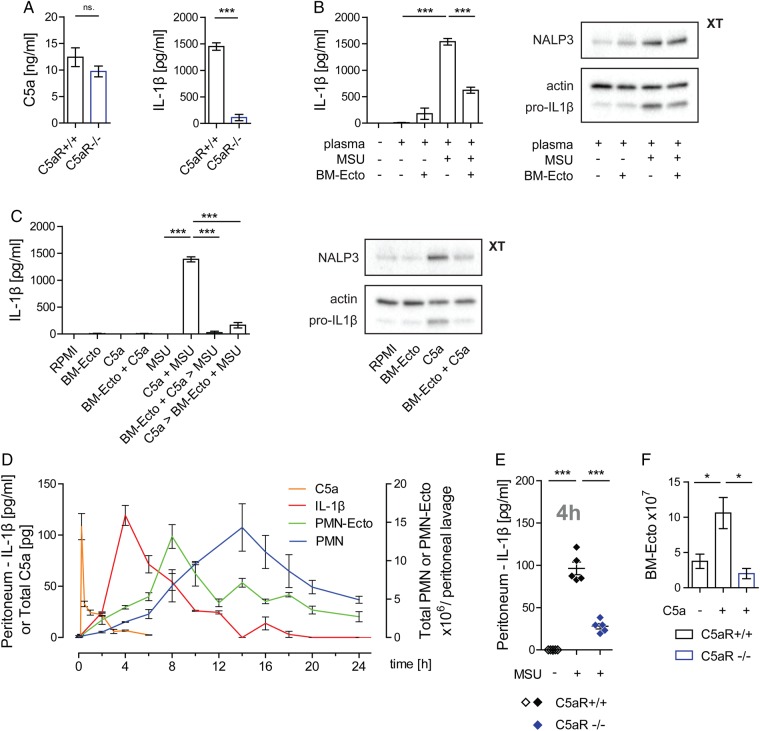
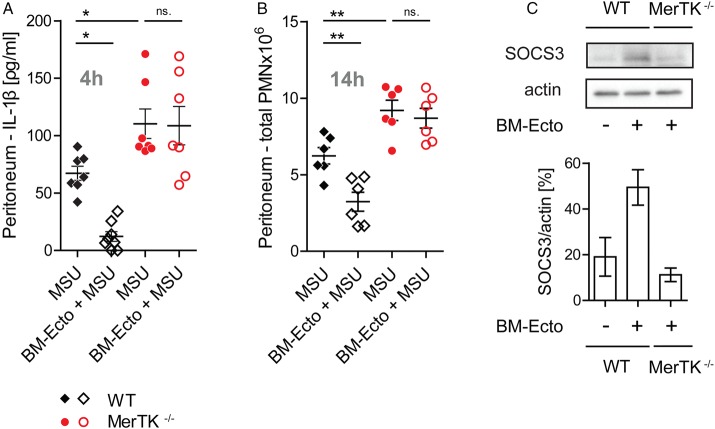
Objectives: Gout is a highly inflammatory but self-limiting joint disease induced by the precipitation of monosodium urate (MSU) crystals. While it is well established that inflammasome activation by MSU mediates acute inflammation, little is known about the mechanism controlling its spontaneous resolution. The aim of this study was to analyse the role of neutrophil-derived microvesicles (PMN-Ecto) in the resolution of acute gout.
Methods: PMN-Ecto were studied in a murine model of MSU-induced peritonitis using C57BL/6, MerTK(-/-) and C5aR(-/-) mice. The peritoneal compartment was assessed for the number of infiltrating neutrophils (PMN), neutrophil microvesicles (PMN-Ecto), cytokines (interleukin-1β, TGFβ) and complement factors (C5a). Human PMN-Ecto were isolated from exudates of patients undergoing an acute gouty attack and functionally tested in vitro.
Results: C5a generated after the injection of MSU primed the inflammasome for IL-1β release. Neutrophils infiltrating the peritoneum in response to C5a released phosphatidylserine (PS)-positive PMN-Ecto early on in the course of inflammation. These PMN-Ecto in turn suppressed C5a priming of the inflammasome and consequently inhibited IL-1β release and neutrophil influx. PMN-Ecto-mediated suppression required surface expression of the PS-receptor MerTK and could be reproduced using PS-expressing liposomes. In addition, ectosomes triggered the release of TGFβ independent of MerTK. TGFβ, however, was not sufficient to control acute MSU-driven inflammation in vivo. Finally, PMN-Ecto from joint aspirates of patients with gouty arthritis had similar anti-inflammatory properties.
Conclusions: PMN-Ecto-mediated control of inflammasome-driven inflammation is a compelling concept of autoregulation initiated early on during PMN activation in gout.
Keywords: Cytokines; Gout; Inflammation.
Published by the BMJ Publishing Group Limited. For permission to use (where not already granted under a licence) please go to http://www.bmj.com/company/products-services/rights-and-licensing/
Figures




Similar articles
-
NLRP3 inflammasome-mediated neutrophil recruitment and hypernociception depend on leukotriene B(4) in a murine model of gout.Arthritis Rheum. 2012 Feb;64(2):474-84. doi: 10.1002/art.33355. Arthritis Rheum. 2012. PMID: 21952942
-
C5a Regulates IL-1β Production and Leukocyte Recruitment in a Murine Model of Monosodium Urate Crystal-Induced Peritonitis.Front Pharmacol. 2017 Jan 23;8:10. doi: 10.3389/fphar.2017.00010. eCollection 2017. Front Pharmacol. 2017. PMID: 28167912 Free PMC article.
-
Resident macrophages initiating and driving inflammation in a monosodium urate monohydrate crystal-induced murine peritoneal model of acute gout.Arthritis Rheum. 2009 Jan;60(1):281-9. doi: 10.1002/art.24185. Arthritis Rheum. 2009. PMID: 19116939
-
Neutrophils, IL-1β, and gout: is there a link?Semin Immunopathol. 2013 Jul;35(4):501-12. doi: 10.1007/s00281-013-0361-0. Epub 2013 Jan 24. Semin Immunopathol. 2013. PMID: 23344781 Review.
-
Mechanisms of spontaneous resolution of acute gouty inflammation.Curr Rheumatol Rep. 2014 Jan;16(1):392. doi: 10.1007/s11926-013-0392-5. Curr Rheumatol Rep. 2014. PMID: 24343224 Review.
Cited by
-
Neutrophils in animal models of autoimmune disease.Semin Immunol. 2016 Apr;28(2):174-86. doi: 10.1016/j.smim.2016.04.001. Epub 2016 Apr 7. Semin Immunol. 2016. PMID: 27067180 Free PMC article. Review.
-
Autoinflammatory Features in Gouty Arthritis.J Clin Med. 2021 Apr 26;10(9):1880. doi: 10.3390/jcm10091880. J Clin Med. 2021. PMID: 33926105 Free PMC article. Review.
-
Alterations in Immune Cell Profiles in the Liver in Diabetes Mellitus: A Systematic Review.Int J Mol Sci. 2025 Apr 24;26(9):4027. doi: 10.3390/ijms26094027. Int J Mol Sci. 2025. PMID: 40362271 Free PMC article.
-
The liver fibrosis niche: Novel insights into the interplay between fibrosis-composing mesenchymal cells, immune cells, endothelial cells, and extracellular matrix.Food Chem Toxicol. 2020 Sep;143:111556. doi: 10.1016/j.fct.2020.111556. Epub 2020 Jul 5. Food Chem Toxicol. 2020. PMID: 32640349 Free PMC article. Review.
-
Neutrophils contribute to spontaneous resolution of liver inflammation and fibrosis via microRNA-223.J Clin Invest. 2019 Jul 11;129(10):4091-4109. doi: 10.1172/JCI122258. J Clin Invest. 2019. PMID: 31295147 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
