

Role of electrostatic interactions for ligand recognition and specificity of peptide transporters
- PMID: 26246134
- PMCID: PMC4527243
- DOI: 10.1186/s12915-015-0167-8
Role of electrostatic interactions for ligand recognition and specificity of peptide transporters
Abstract
Background: Peptide transporters are membrane proteins that mediate the cellular uptake of di- and tripeptides, and of peptidomimetic drugs such as β-lactam antibiotics, antiviral drugs and antineoplastic agents. In spite of their high physiological and pharmaceutical importance, the molecular recognition by these transporters of the amino acid side chains of short peptides and thus the mechanisms for substrate binding and specificity are far from being understood.
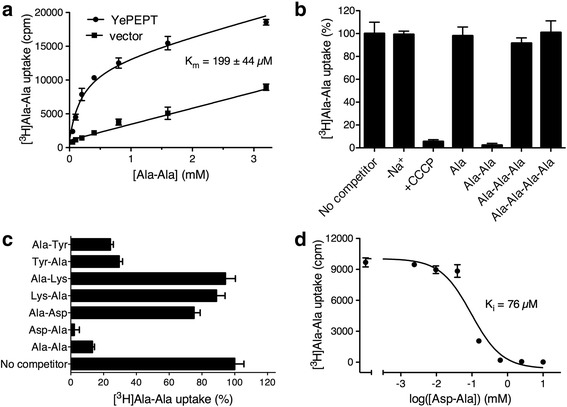
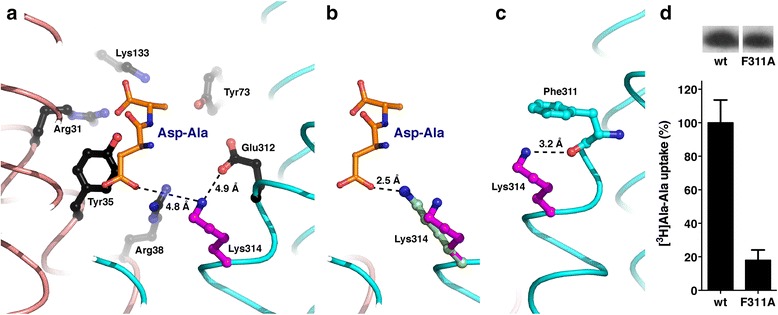
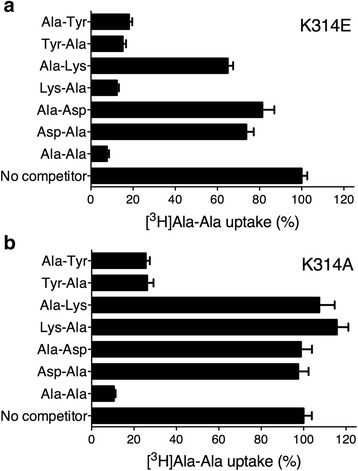
Results: The X-ray crystal structure of the peptide transporter YePEPT from the bacterium Yersinia enterocolitica together with functional studies have unveiled the molecular bases for recognition, binding and specificity of dipeptides with a charged amino acid residue at the N-terminal position. In wild-type YePEPT, the significant specificity for the dipeptides Asp-Ala and Glu-Ala is defined by electrostatic interaction between the in the structure identified positively charged Lys314 and the negatively charged amino acid side chain of these dipeptides. Mutagenesis of Lys314 into the negatively charged residue Glu allowed tuning of the substrate specificity of YePEPT for the positively charged dipeptide Lys-Ala. Importantly, molecular insights acquired from the prokaryotic peptide transporter YePEPT combined with mutagenesis and functional uptake studies with human PEPT1 expressed in Xenopus oocytes also allowed tuning of human PEPT1's substrate specificity, thus improving our understanding of substrate recognition and specificity of this physiologically and pharmaceutically important peptide transporter.
Conclusion: This study provides the molecular bases for recognition, binding and specificity of peptide transporters for dipeptides with a charged amino acid residue at the N-terminal position.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
