

Endogenous DUX4 expression in FSHD myotubes is sufficient to cause cell death and disrupts RNA splicing and cell migration pathways
- PMID: 26246499
- PMCID: PMC4581613
- DOI: 10.1093/hmg/ddv315
Endogenous DUX4 expression in FSHD myotubes is sufficient to cause cell death and disrupts RNA splicing and cell migration pathways
Abstract
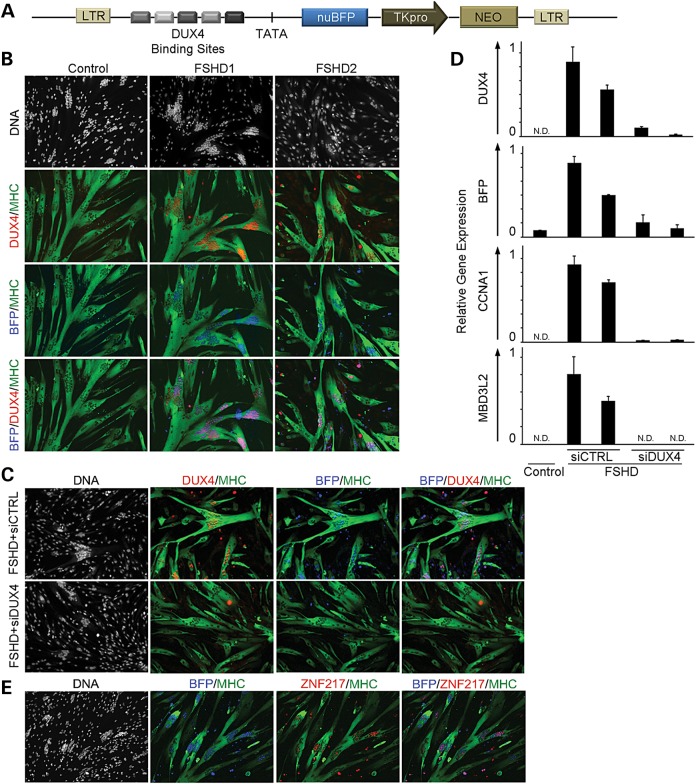
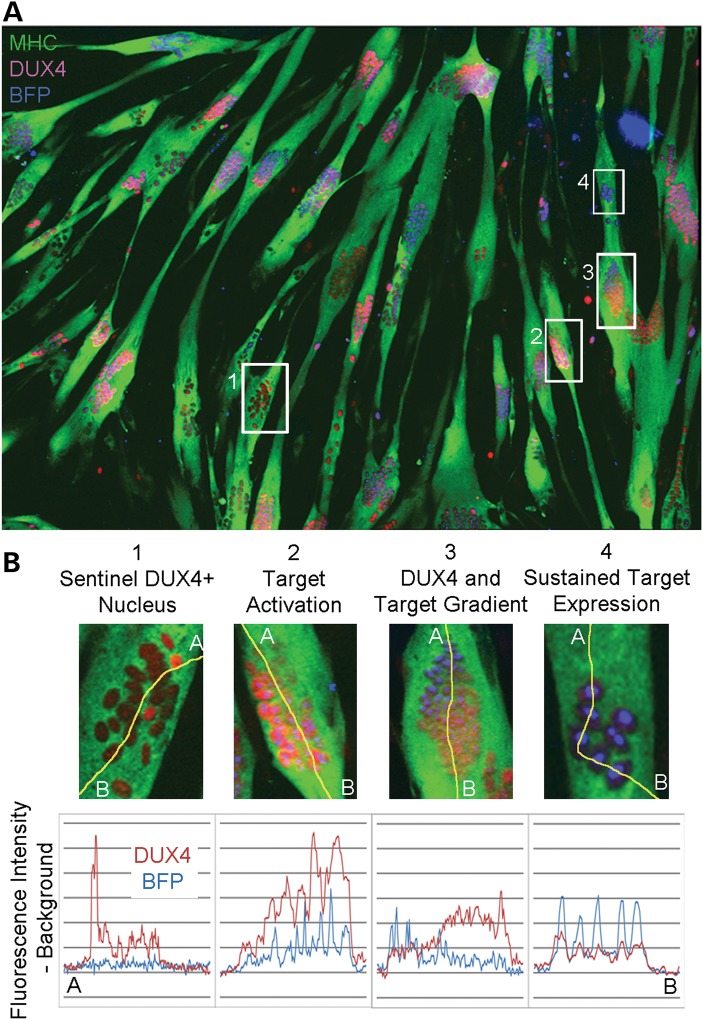
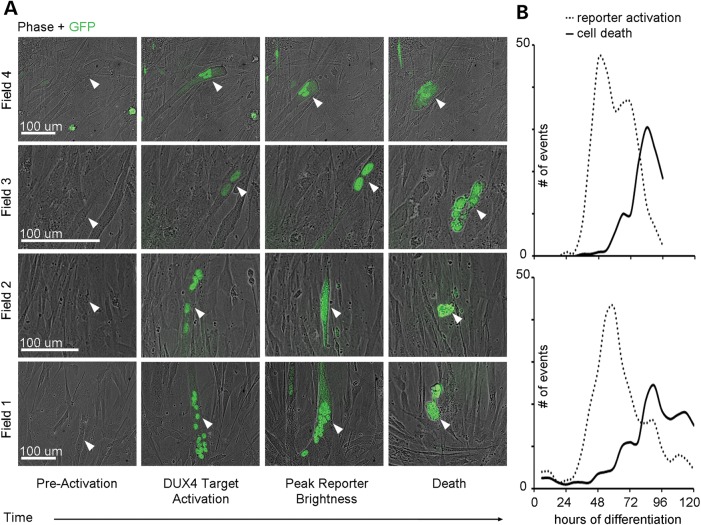
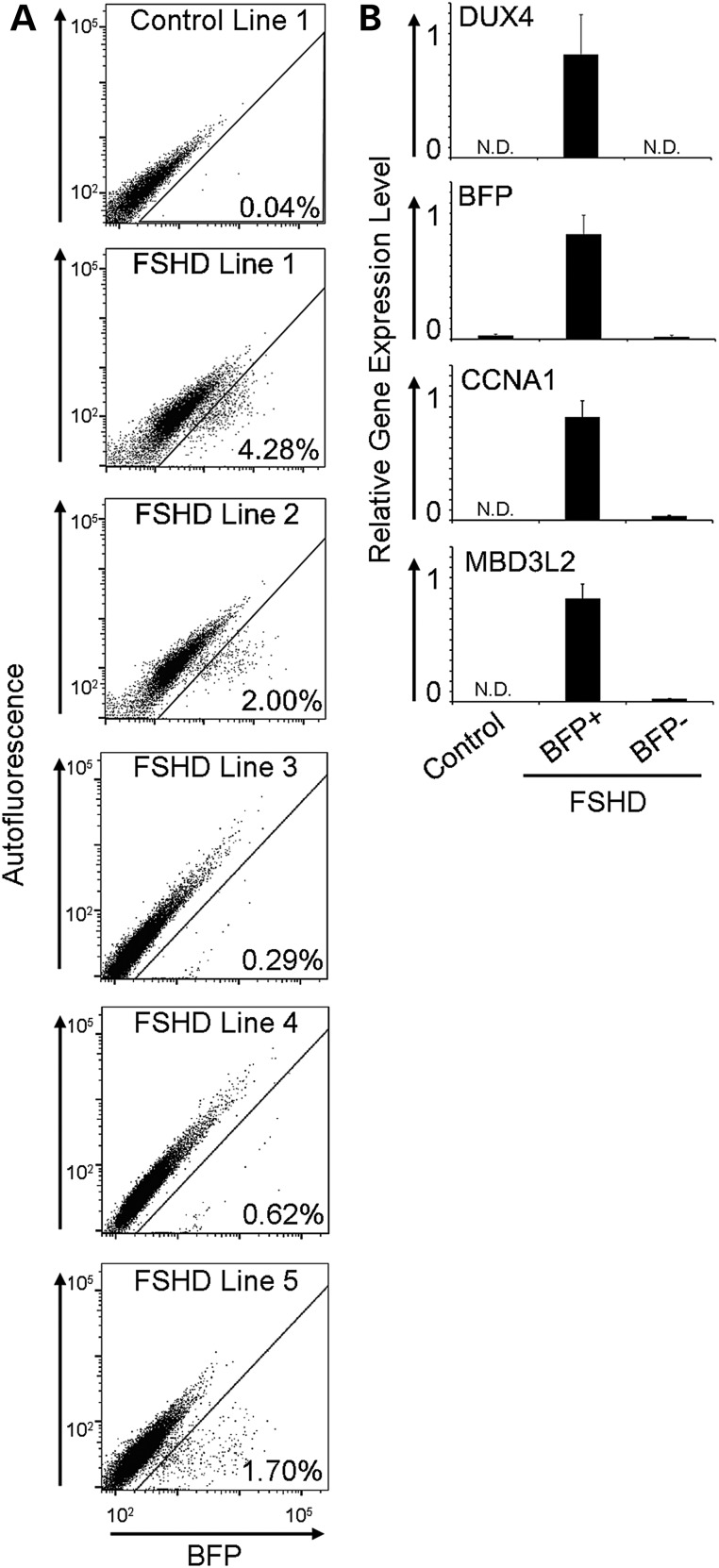
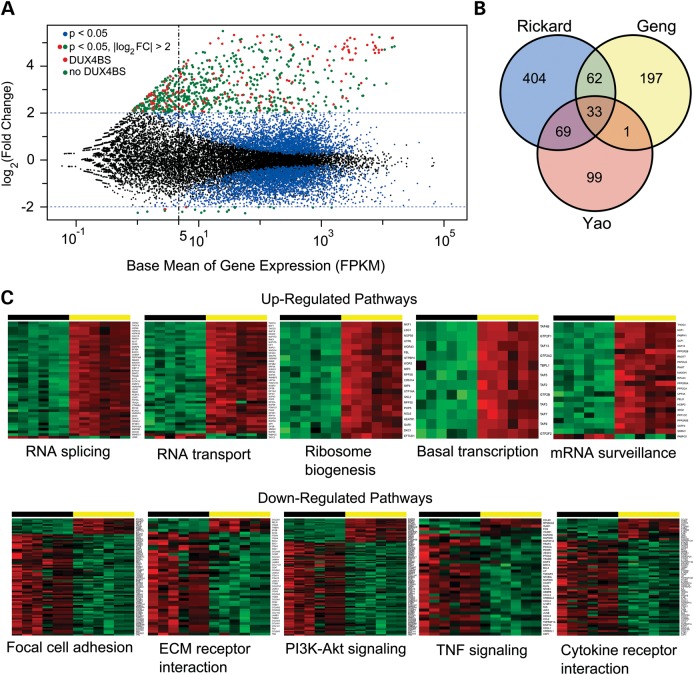
Facioscapulohumeral muscular dystrophy (FSHD) is caused by chromatin relaxation that results in aberrant expression of the transcription factor Double Homeobox 4 (DUX4). DUX4 protein is present in a small subset of FSHD muscle cells, making its detection and analysis of its effects historically difficult. Using a DUX4-activated reporter, we demonstrate the burst expression pattern of endogenous DUX4, its method of signal amplification in the unique shared cytoplasm of the myotube, and FSHD cell death that depends on its activation. Transcriptome analysis of DUX4-expressing cells revealed that DUX4 activation disrupts RNA metabolism including RNA splicing, surveillance and transport pathways. Cell signaling, polarity and migration pathways were also disrupted. Thus, DUX4 expression is sufficient for myocyte death, and these findings suggest mechanistic links between DUX4 expression and cell migration, supporting recent descriptions of phenotypic similarities between FSHD and an FSHD-like condition caused by FAT1 mutations.
© The Author 2015. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Taylor D.A., Carroll J.E., Smith M.E., Johnson M.O., Johnston G.P., Brooke M.H. (1982) Facioscapulohumeral dystrophy associated with hearing loss and Coats syndrome. Ann. Neurol., 12, 395–398. - PubMed
-
- Gurwin E.B., Fitzsimons R.B., Sehmi K.S., Bird A.C. (1985) Retinal telangiectasis in facioscapulohumeral muscular dystrophy with deafness. Arch. Ophthalmol., 103, 1695–1700. - PubMed
-
- van Overveld P.G., Lemmers R.J., Sandkuijl L.A., Enthoven L., Winokur S.T., Bakels F., Padberg G.W., van Ommen G.J., Frants R.R., van der Maarel S.M. (2003) Hypomethylation of D4Z4 in 4q-linked and non-4q-linked facioscapulohumeral muscular dystrophy. Nat. Genet., 35, 315–317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
