

Replication stress in early S phase generates apparent micronuclei and chromosome rearrangement in fission yeast
- PMID: 26246602
- PMCID: PMC4591689
- DOI: 10.1091/mbc.E15-05-0318
Replication stress in early S phase generates apparent micronuclei and chromosome rearrangement in fission yeast
Abstract
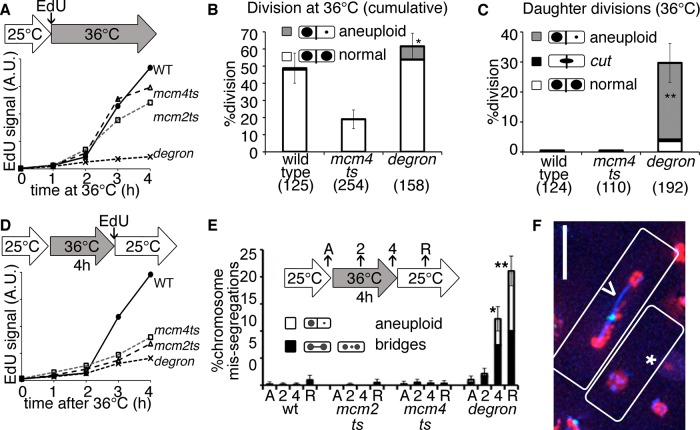
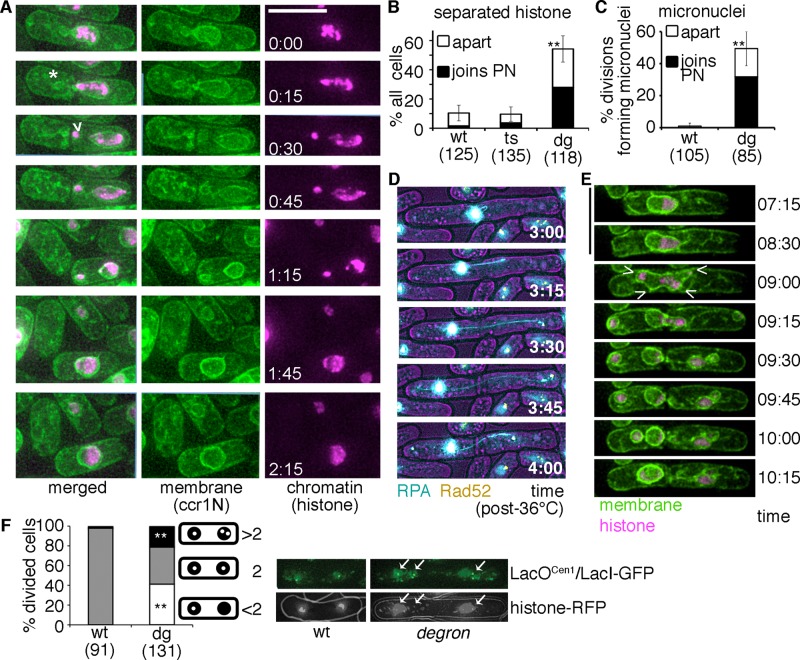
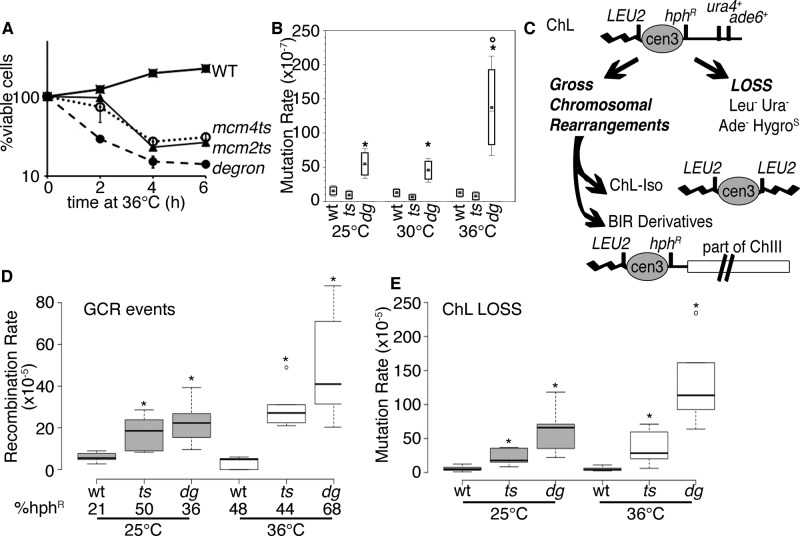
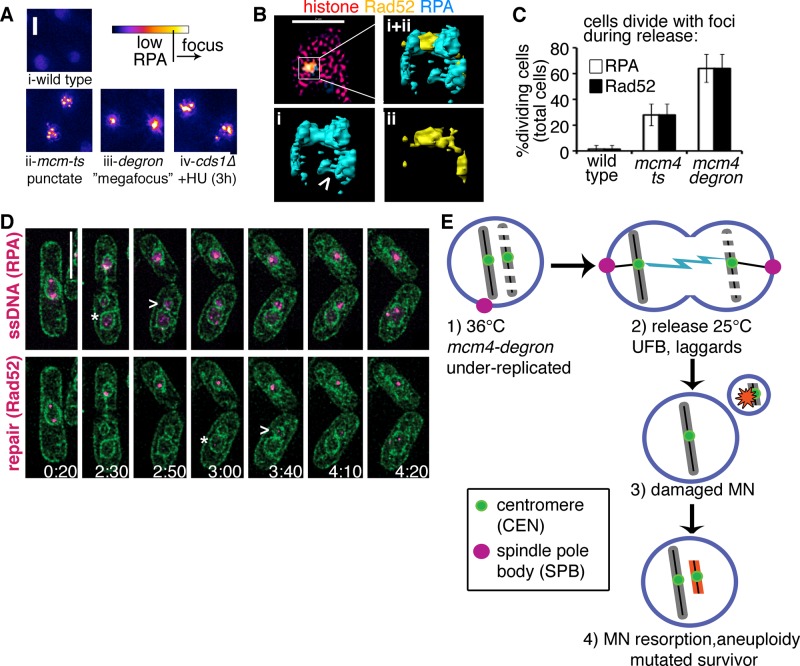
DNA replication stress causes genome mutations, rearrangements, and chromosome missegregation, which are implicated in cancer. We analyze a fission yeast mutant that is unable to complete S phase due to a defective subunit of the MCM helicase. Despite underreplicated and damaged DNA, these cells evade the G2 damage checkpoint to form ultrafine bridges, fragmented centromeres, and uneven chromosome segregations that resembles micronuclei. These micronuclei retain DNA damage markers and frequently rejoin with the parent nucleus. Surviving cells show an increased rate of mutation and chromosome rearrangement. This first report of micronucleus-like segregation in a yeast replication mutant establishes underreplication as an important factor contributing to checkpoint escape, abnormal chromosome segregation, and chromosome instability.
© 2015 Sabatinos et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
-
- Bartkova J, Horejsi Z, Koed K, Kramer A, Tort F, Zieger K, Guldberg P, Sehested M, Nesland JM, Lukas C, et al. DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature. 2005;434:864–870. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
