

Developmental alterations in centrosome integrity contribute to the post-mitotic state of mammalian cardiomyocytes
- PMID: 26247711
- PMCID: PMC4541494
- DOI: 10.7554/eLife.05563
Developmental alterations in centrosome integrity contribute to the post-mitotic state of mammalian cardiomyocytes
Abstract
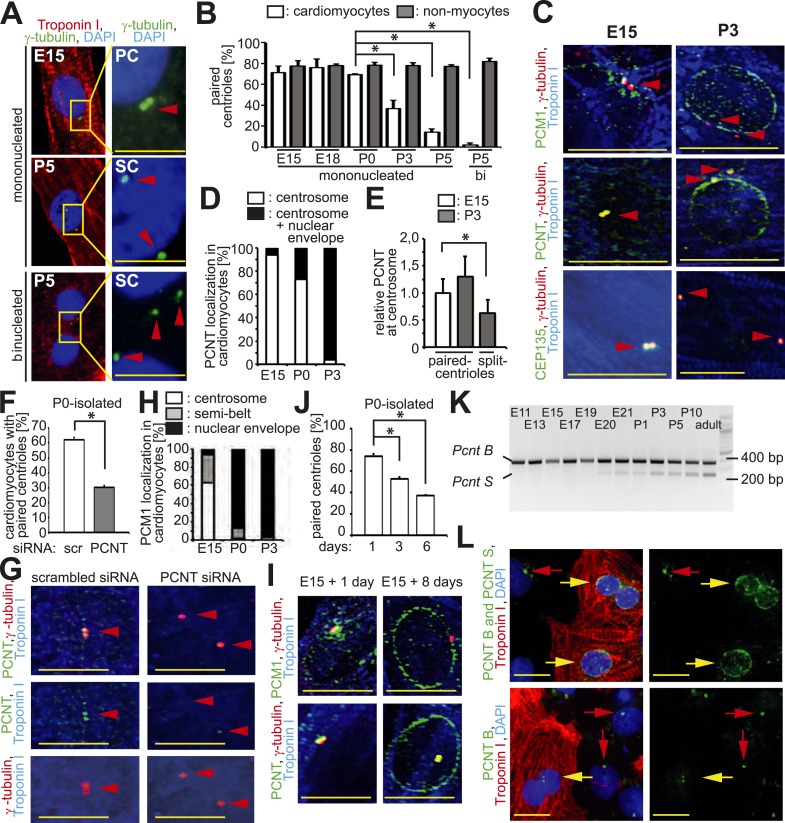
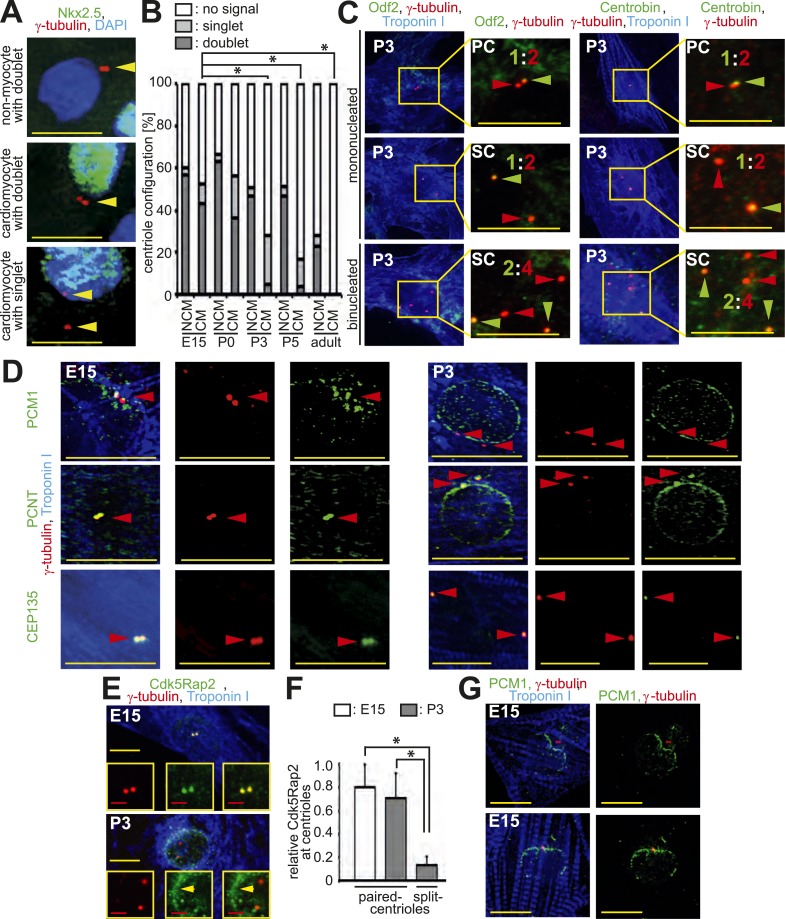
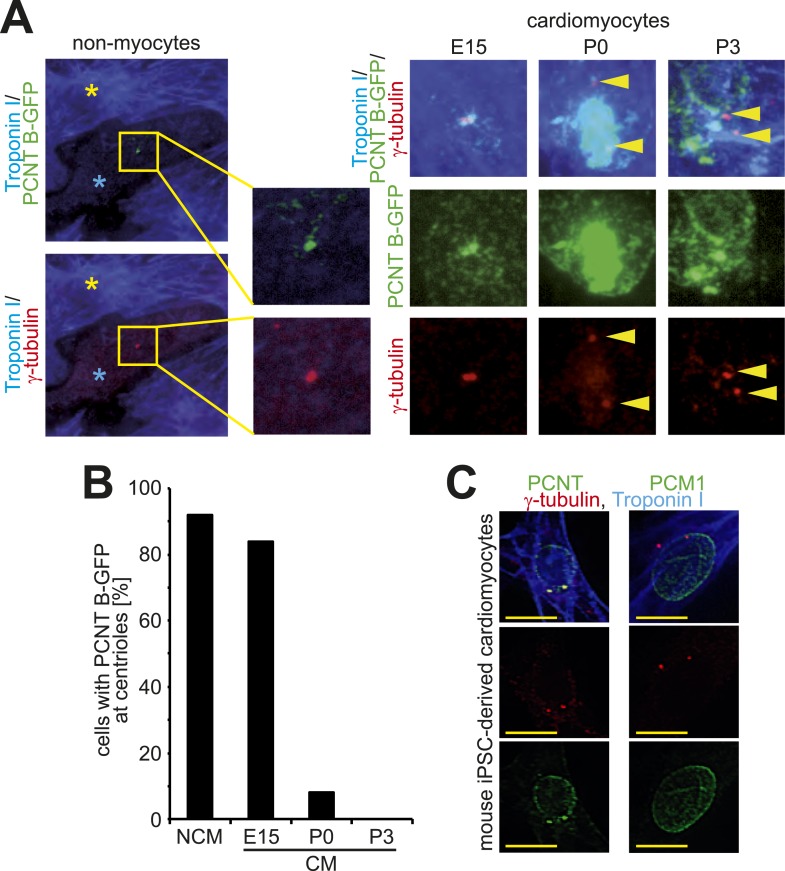
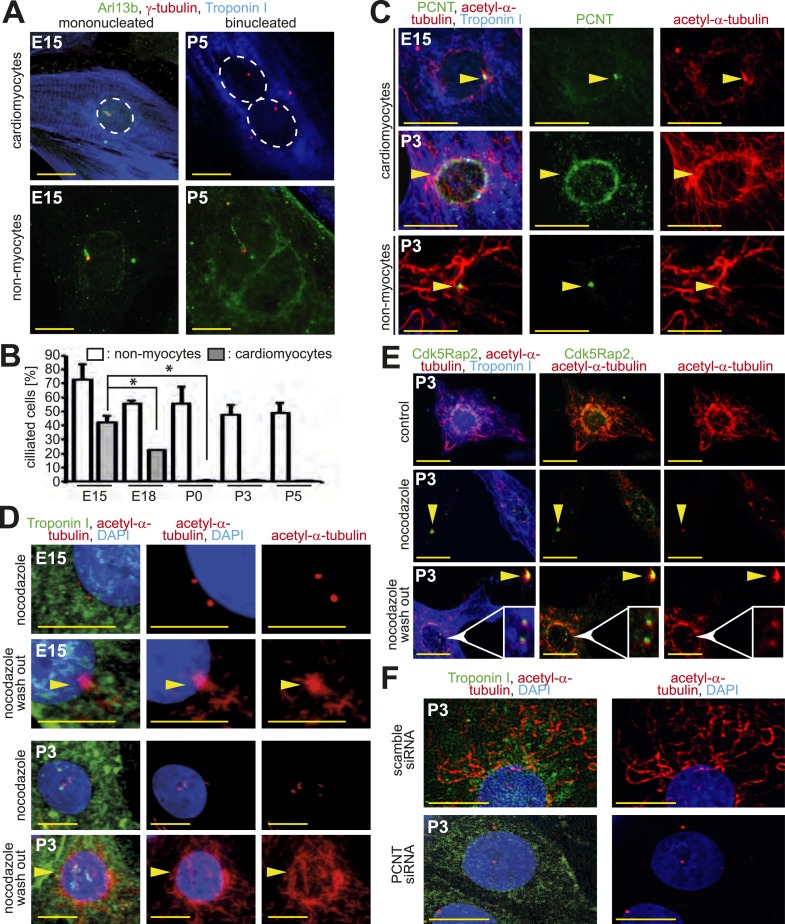
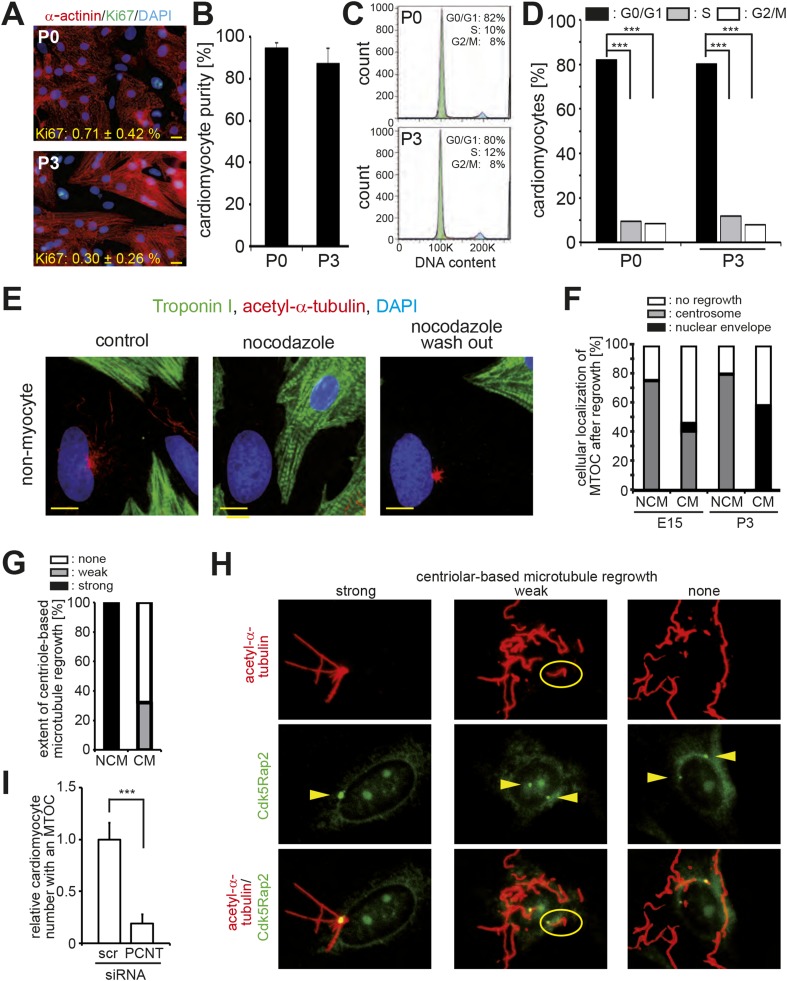
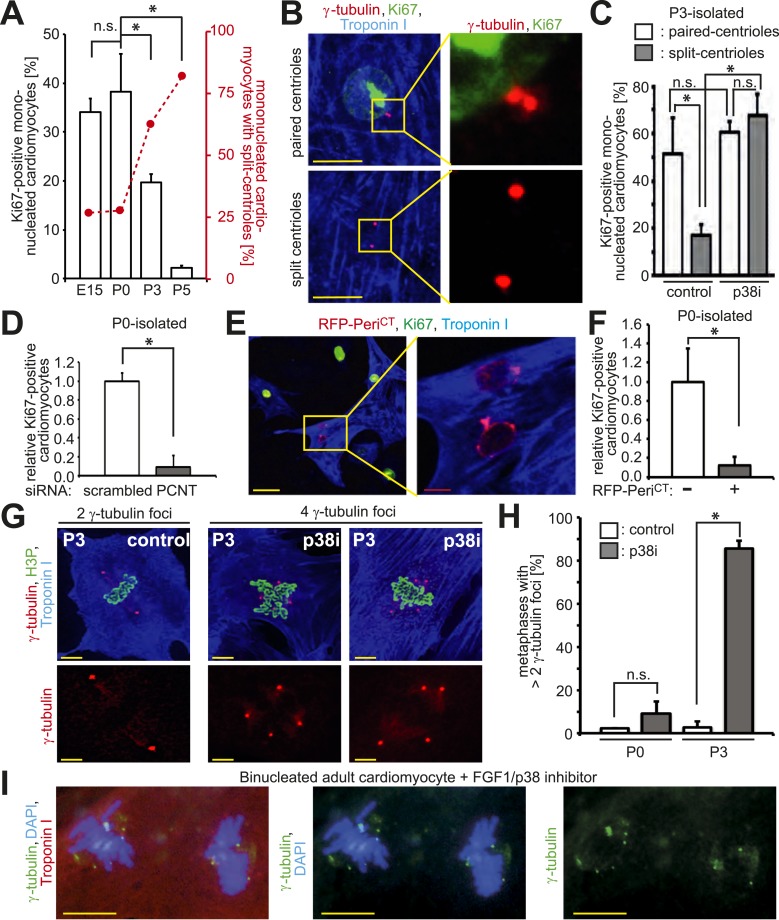
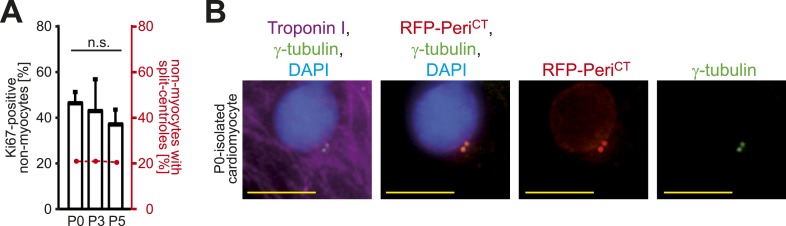
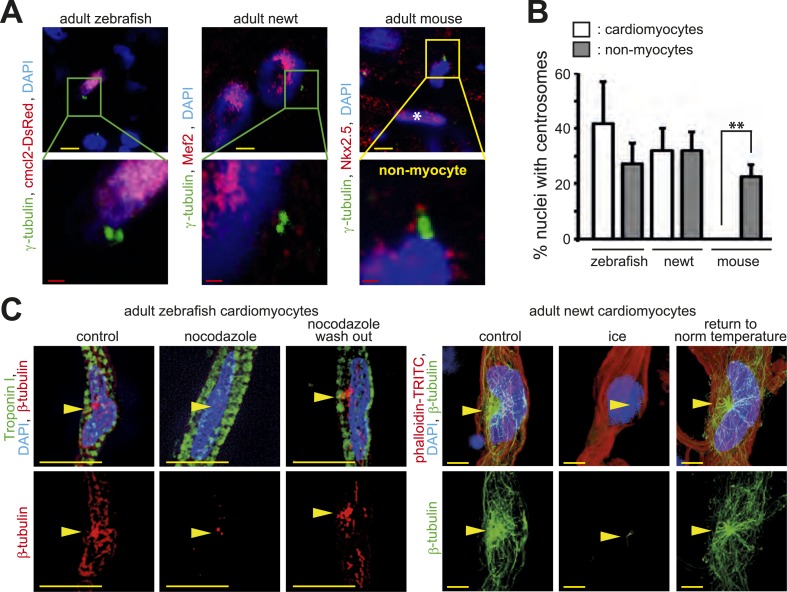
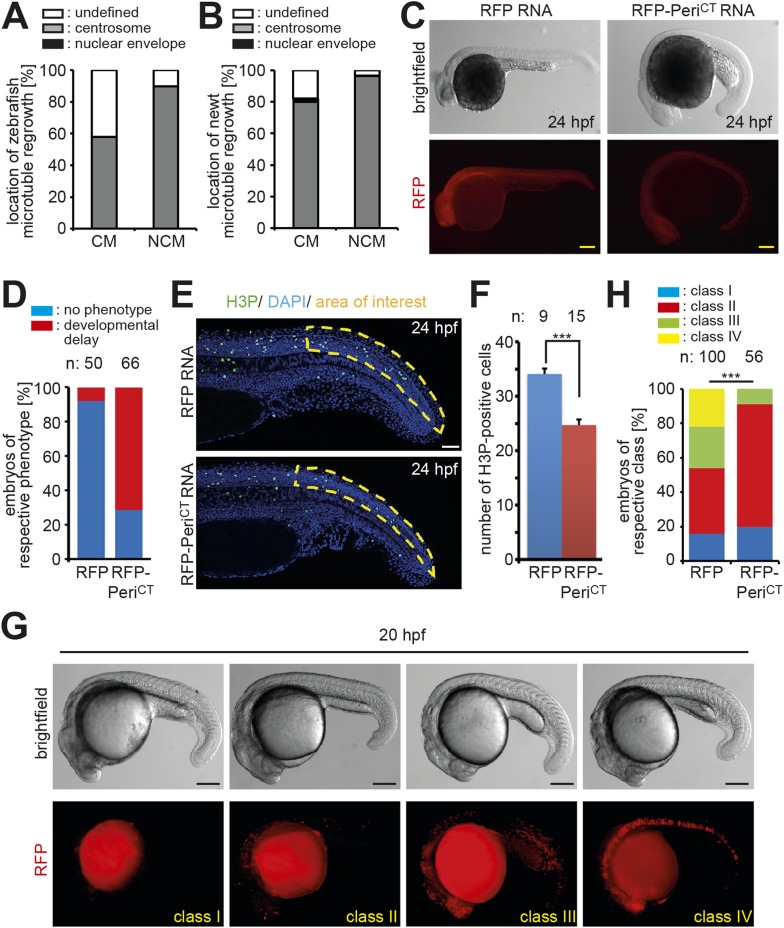
Mammalian cardiomyocytes become post-mitotic shortly after birth. Understanding how this occurs is highly relevant to cardiac regenerative therapy. Yet, how cardiomyocytes achieve and maintain a post-mitotic state is unknown. Here, we show that cardiomyocyte centrosome integrity is lost shortly after birth. This is coupled with relocalization of various centrosome proteins to the nuclear envelope. Consequently, postnatal cardiomyocytes are unable to undergo ciliogenesis and the nuclear envelope adopts the function as cellular microtubule organizing center. Loss of centrosome integrity is associated with, and can promote, cardiomyocyte G0/G1 cell cycle arrest suggesting that centrosome disassembly is developmentally utilized to achieve the post-mitotic state in mammalian cardiomyocytes. Adult cardiomyocytes of zebrafish and newt, which are able to proliferate, maintain centrosome integrity. Collectively, our data provide a novel mechanism underlying the post-mitotic state of mammalian cardiomyocytes as well as a potential explanation for why zebrafish and newts, but not mammals, can regenerate their heart.
Keywords: MTOC; cardiomyocyte; cardiomyocyte proliferation; cell biology; centrosome; developmental biology; heart regeneration; mouse; newt; primary cilium; rat; stem cells; terminal differentiation; zebrafish.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
