

Relationship between serum response factor and androgen receptor in prostate cancer
- PMID: 26250344
- PMCID: PMC4579008
- DOI: 10.1002/pros.23051
Relationship between serum response factor and androgen receptor in prostate cancer
Abstract
Background: Serum response factor (SRF) is an important transcription factor in castrate-resistant prostate cancer (CRPC). Since CRPC is associated with androgen receptor (AR) hypersensitivity, we investigated the relationship between SRF and AR.
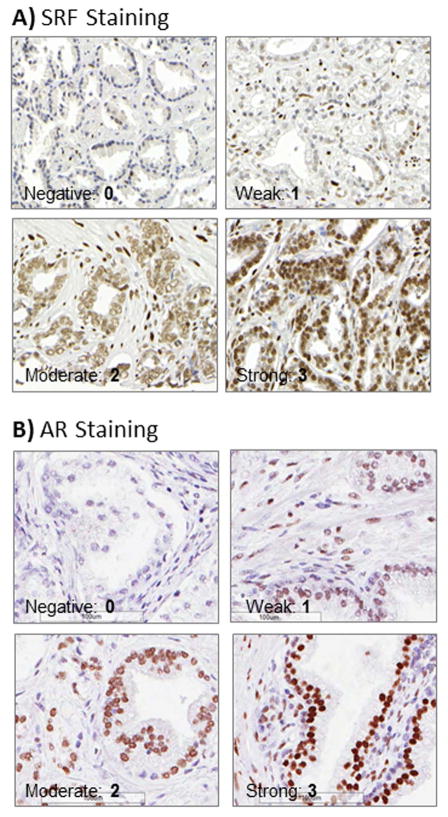
Materials and methods: Transcriptional activity was assessed by luciferase assay. Cell proliferation was measured by MTT and flow cytometry. Protein expression in patients was assessed by immunohistochemistry.
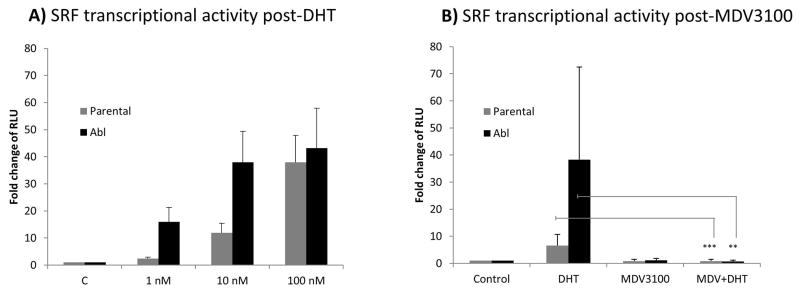
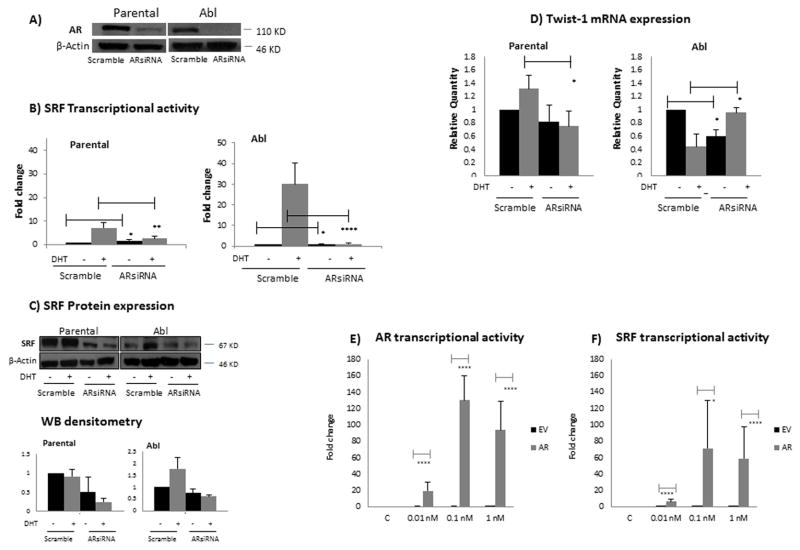
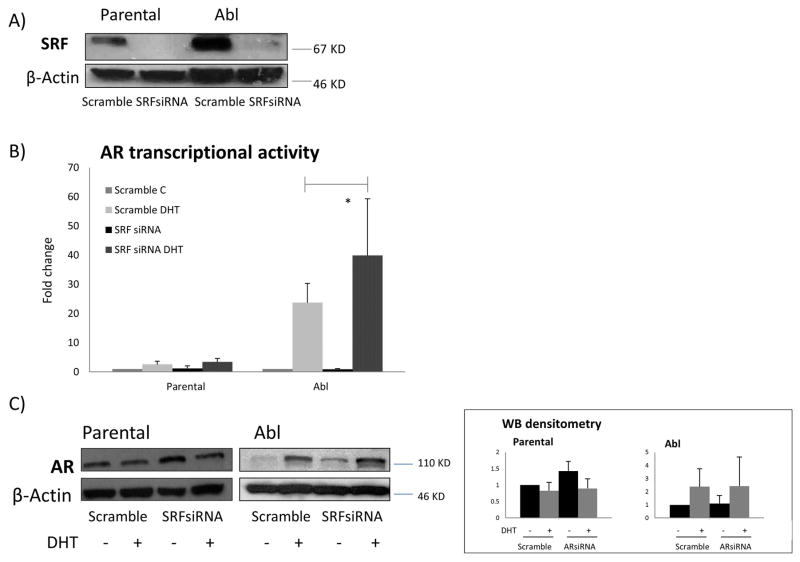
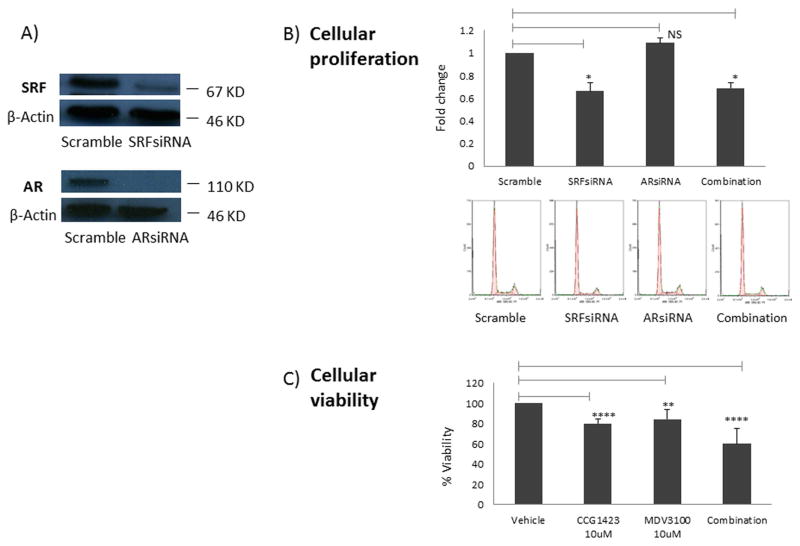
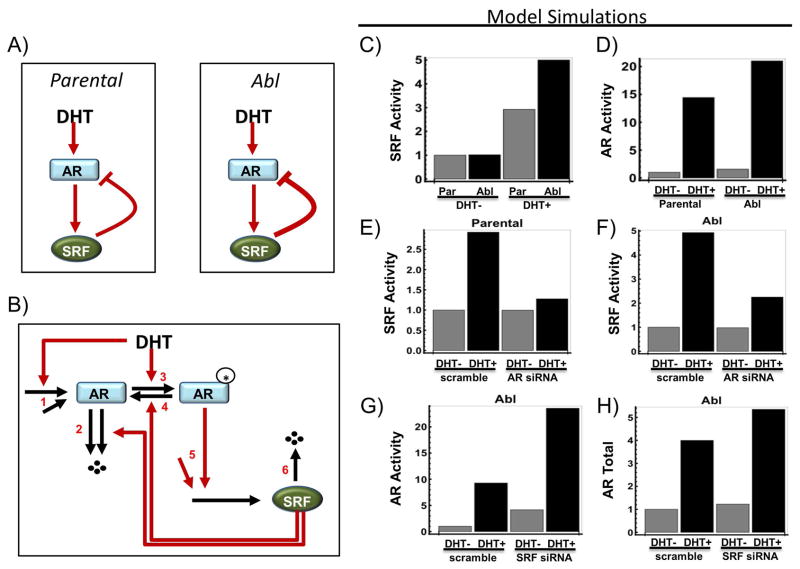
Results: To investigate AR involvement in SRF response to androgen, AR expression was down-regulated using siRNA. This resulted in the abrogation of SRF induction post-DHT. Moreover, DHT stimulation failed to induce SRF transcriptional activity in AR-negative PC346 DCC cells, which was only restored following AR over-expression. Next, SRF expression was down-regulated by siRNA, resulting in AR increased transcriptional activity in castrate-resistant LNCaP Abl cells but not in the parental LNCaP. This negative feedback loop in the resistant cells was confirmed by immunohistochemistry which showed a negative correlation between AR and SRF expression in CRPC bone metastases and a positive correlation in androgen-naïve prostatectomies. Cell proliferation was next assessed following SRF inhibition, demonstrating that SRF inhibition is more effective than AR inhibition in castrate-resistant cells.
Conclusion: Our data support SRF as a promising therapeutic target in combination with current treatments.
Keywords: androgen receptor; castrate-resistant prostate cancer; serum response factor.
© 2015 Wiley Periodicals, Inc.
Figures
References
-
- Prencipe M, Madden SF, O’Neill A, O’Hurley Gillian, Culhane Aedin, O’Connor Darran, Klocker H, Kay EW, Gallagher WM, Watson WR. Identification of transcription factors associated with castration-resistance: is the Serum Responsive Factor a potential therapeutic target? Prostate. 2013;73(7):743–53. - PubMed
-
- Vickers ER, Kasza A, Kurnaz IA, Seifert A, Zeef LA, O’donnell A, Hayes A, Sharrocks AD. Ternary complex factor-serum response factor complex-regulated gene activity is required for cellular proliferation and inhibition of apoptotic cell death. Molecular and cellular biology. 2004;24(23):10340–51. - PMC - PubMed
-
- Heemers HV, Regan KM, Dehm SM, Tindall DJ. Androgen induction of the androgen receptor coactivator four and a half LIM domain protein-2: evidence for a role for serum response factor in prostate cancer. Cancer Res. 2007;67(21):10592–9. - PubMed
-
- O’Hurley G, Prencipe M, Lundon D, O’Neill A, Boyce S, O’Grady A, Gallagher WM, Morrissey C, Kay EW, Watson W. The analysis of Serum Response Factor expression in bone and soft tissue Prostate Cancer metastases. Prostate. 2014;74:306–313. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
