

Astrocyte reactivity after brain injury-: The role of galectins 1 and 3
- PMID: 26250529
- PMCID: PMC5042059
- DOI: 10.1002/glia.22898
Astrocyte reactivity after brain injury-: The role of galectins 1 and 3
Abstract
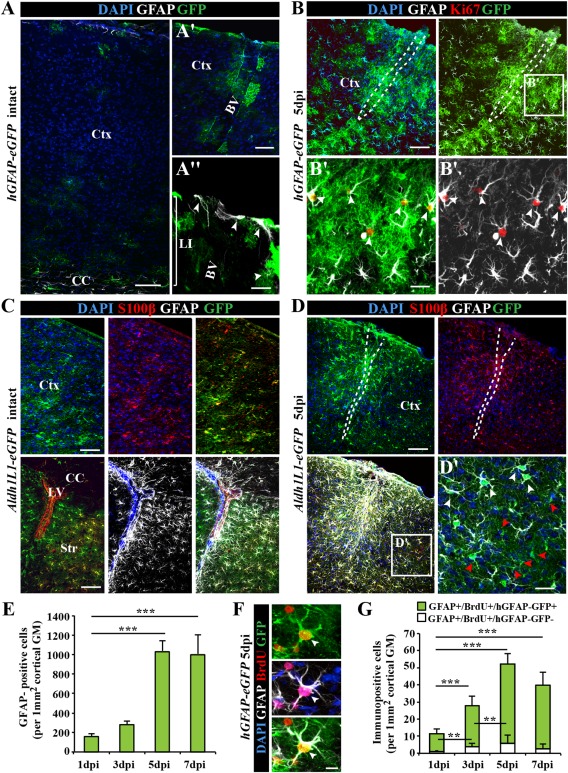
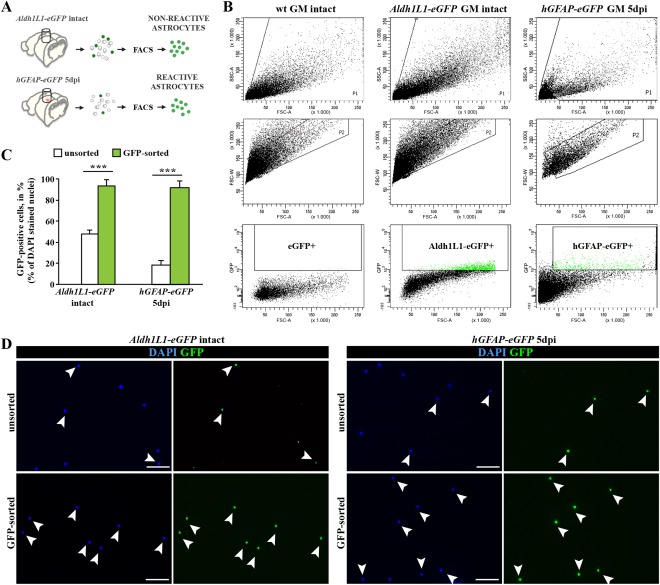
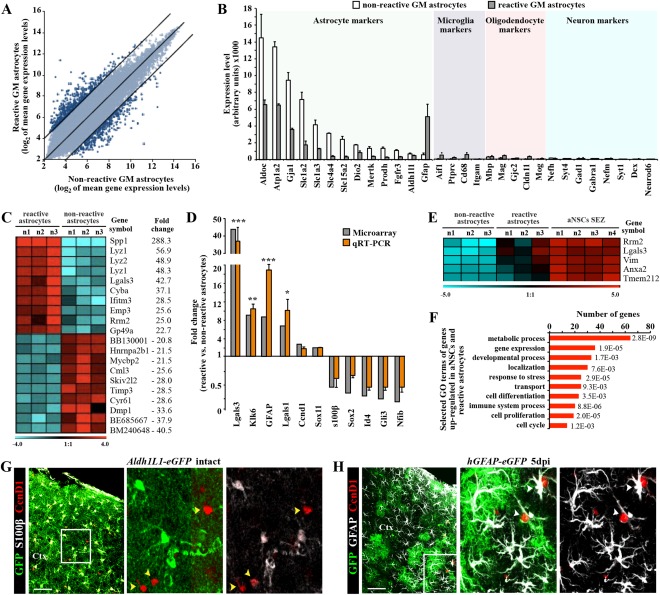
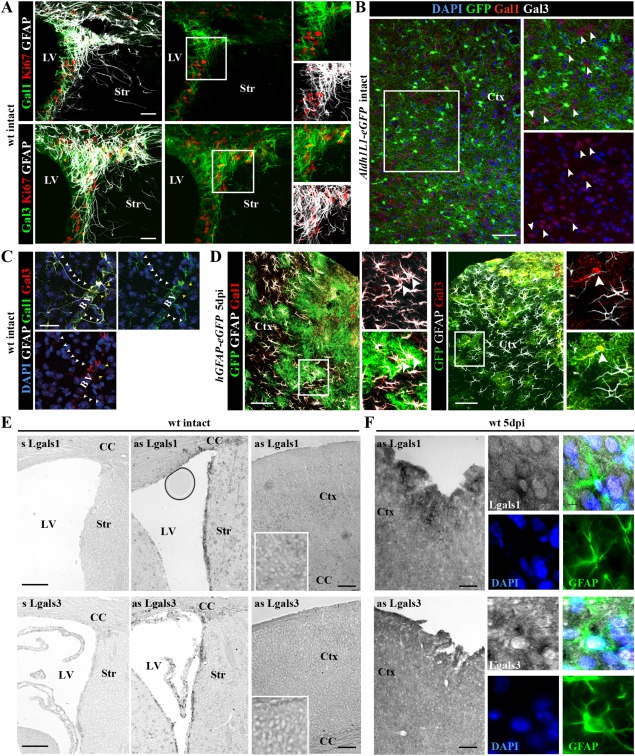
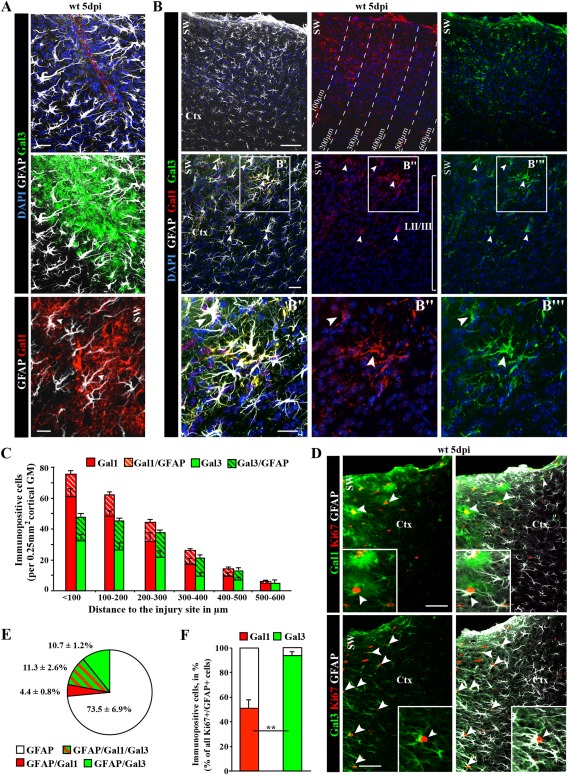
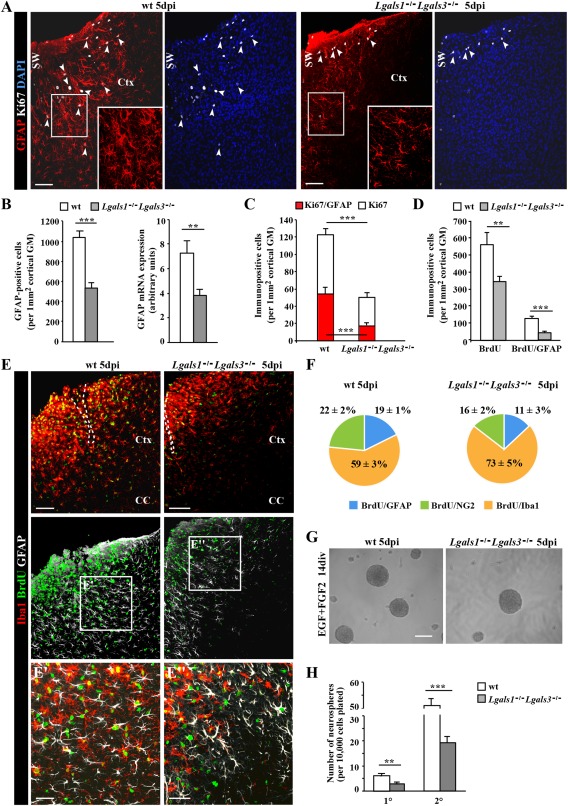
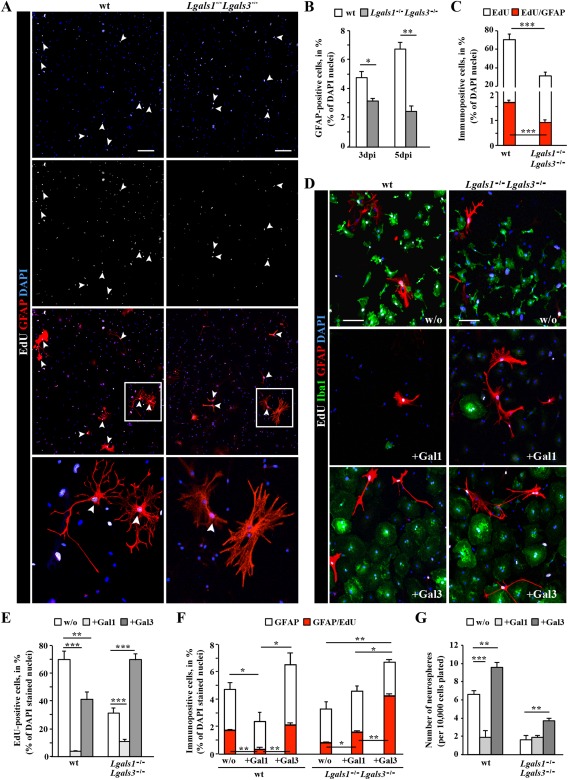
Astrocytes react to brain injury in a heterogeneous manner with only a subset resuming proliferation and acquiring stem cell properties in vitro. In order to identify novel regulators of this subset, we performed genomewide expression analysis of reactive astrocytes isolated 5 days after stab wound injury from the gray matter of adult mouse cerebral cortex. The expression pattern was compared with astrocytes from intact cortex and adult neural stem cells (NSCs) isolated from the subependymal zone (SEZ). These comparisons revealed a set of genes expressed at higher levels in both endogenous NSCs and reactive astrocytes, including two lectins-Galectins 1 and 3. These results and the pattern of Galectin expression in the lesioned brain led us to examine the functional significance of these lectins in brains of mice lacking Galectins 1 and 3. Following stab wound injury, astrocyte reactivity including glial fibrillary acidic protein expression, proliferation and neurosphere-forming capacity were found significantly reduced in mutant animals. This phenotype could be recapitulated in vitro and was fully rescued by addition of Galectin 3, but not of Galectin 1. Thus, Galectins 1 and 3 play key roles in regulating the proliferative and NSC potential of a subset of reactive astrocytes.
Keywords: genomewide analysis; glia proliferation; neurosphere.
© 2015 The Authors. Glia Published by Wiley Periodicals, Inc.
Figures
Similar articles
-
Inflammation Promotes a Conversion of Astrocytes into Neural Progenitor Cells via NF-κB Activation.Mol Neurobiol. 2016 Oct;53(8):5041-55. doi: 10.1007/s12035-015-9428-3. Epub 2015 Sep 17. Mol Neurobiol. 2016. PMID: 26381429 Free PMC article.
-
FABP7 expression in normal and stab-injured brain cortex and its role in astrocyte proliferation.Histochem Cell Biol. 2011 Nov;136(5):501-13. doi: 10.1007/s00418-011-0865-4. Epub 2011 Sep 22. Histochem Cell Biol. 2011. PMID: 21938553 Free PMC article.
-
Regulation of adult neural progenitor cells by Galectin-1/beta1 Integrin interaction.J Neurochem. 2010 Jun;113(6):1516-24. doi: 10.1111/j.1471-4159.2010.06712.x. Epub 2010 Mar 29. J Neurochem. 2010. PMID: 20367753
-
Thermodynamic binding studies of galectin-1, -3 and -7.Glycoconj J. 2002;19(7-9):459-65. doi: 10.1023/B:GLYC.0000014075.62724.d0. Glycoconj J. 2002. PMID: 14758069 Review.
-
Expression of galectins in cancer: a critical review.Glycoconj J. 2002;19(7-9):537-42. doi: 10.1023/B:GLYC.0000014083.48508.6a. Glycoconj J. 2002. PMID: 14758077 Review.
Cited by
-
Drebrin controls scar formation and astrocyte reactivity upon traumatic brain injury by regulating membrane trafficking.Nat Commun. 2021 Mar 5;12(1):1490. doi: 10.1038/s41467-021-21662-x. Nat Commun. 2021. PMID: 33674568 Free PMC article.
-
The emerging role of galectins in (re)myelination and its potential for developing new approaches to treat multiple sclerosis.Cell Mol Life Sci. 2020 Apr;77(7):1289-1317. doi: 10.1007/s00018-019-03327-7. Epub 2019 Oct 18. Cell Mol Life Sci. 2020. PMID: 31628495 Free PMC article. Review.
-
Circulating extracellular vesicles in facilitated stroke recovery via MiR-451-5p/MIF and MiR-451-5p/CCND1 axes.J Cereb Blood Flow Metab. 2025 Jul 28:271678X251361247. doi: 10.1177/0271678X251361247. Online ahead of print. J Cereb Blood Flow Metab. 2025. PMID: 40719611 Free PMC article.
-
Microglia control small vessel calcification via TREM2.Sci Adv. 2021 Feb 26;7(9):eabc4898. doi: 10.1126/sciadv.abc4898. Print 2021 Feb. Sci Adv. 2021. PMID: 33637522 Free PMC article.
-
Transdifferentiating Astrocytes Into Neurons Using ASCL1 Functionalized With a Novel Intracellular Protein Delivery Technology.Front Bioeng Biotechnol. 2018 Nov 21;6:173. doi: 10.3389/fbioe.2018.00173. eCollection 2018. Front Bioeng Biotechnol. 2018. PMID: 30525033 Free PMC article.
References
-
- Astorgues‐Xerri L, Riveiro ME, Tijeras‐Raballand A, Serova M, Neuzillet C, Albert S, Raymond E, Faivre S. 2014. Unraveling galectin‐1 as a novel therapeutic target for cancer. Cancer Treat Rev 40:307–319. - PubMed
-
- Bardehle S, Krüger M, Buggenthin F, Schwausch J, Ninkovic J, Clevers H, Snippert HJ, Theis FJ, Meyer‐Luehmann M, Bechmann I, Dimou L, Götz M. 2013. Live imaging of astrocyte responses to acute injury reveals selective juxtavascular proliferation. Nat Neurosci 16:580–586. - PubMed
-
- Beckervordersandforth R, Tripathi P, Ninkovic J, Bayam E, Lepier A, Stempfhuber B, Kirchhoff F, Hirrlinger J, Haslinger A, Lie DC, Beckers J, Yoder B, Irmler M, Götz M. 2010. In vivo fate mapping and expression analysis reveals molecular hallmarks of prospectively isolated adult neural stem cells. Cell Stem Cell 7:744–758. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
