

THZ1 Reveals Roles for Cdk7 in Co-transcriptional Capping and Pausing
- PMID: 26257281
- PMCID: PMC4546572
- DOI: 10.1016/j.molcel.2015.06.032
THZ1 Reveals Roles for Cdk7 in Co-transcriptional Capping and Pausing
Abstract
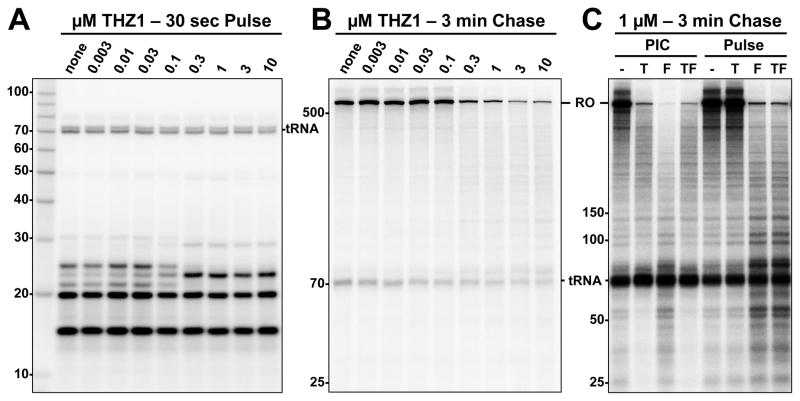
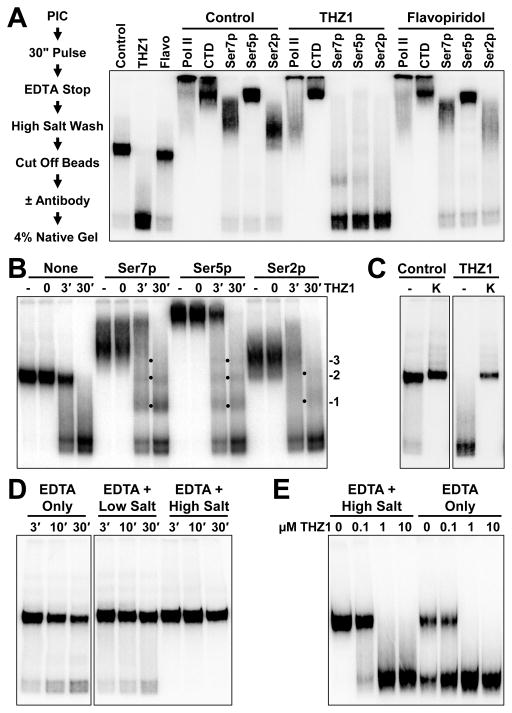
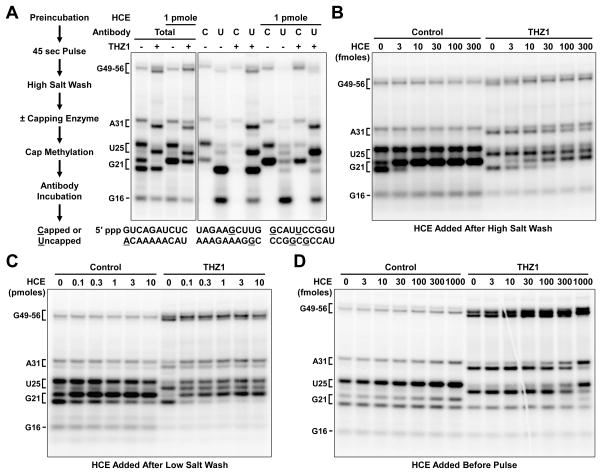
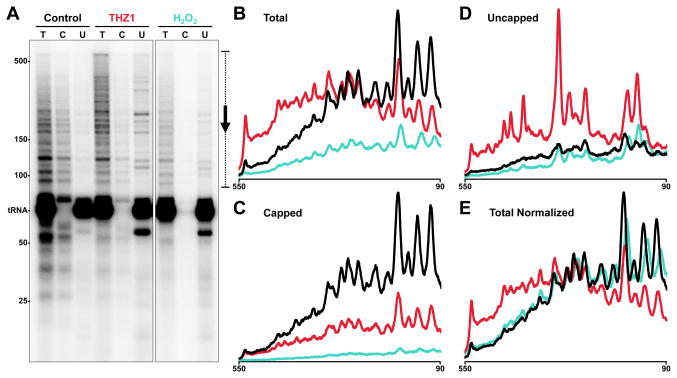
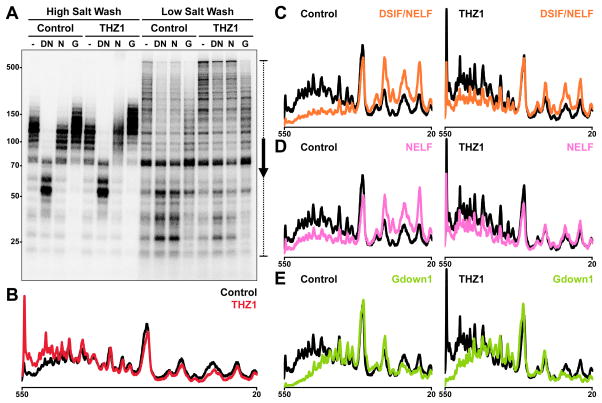
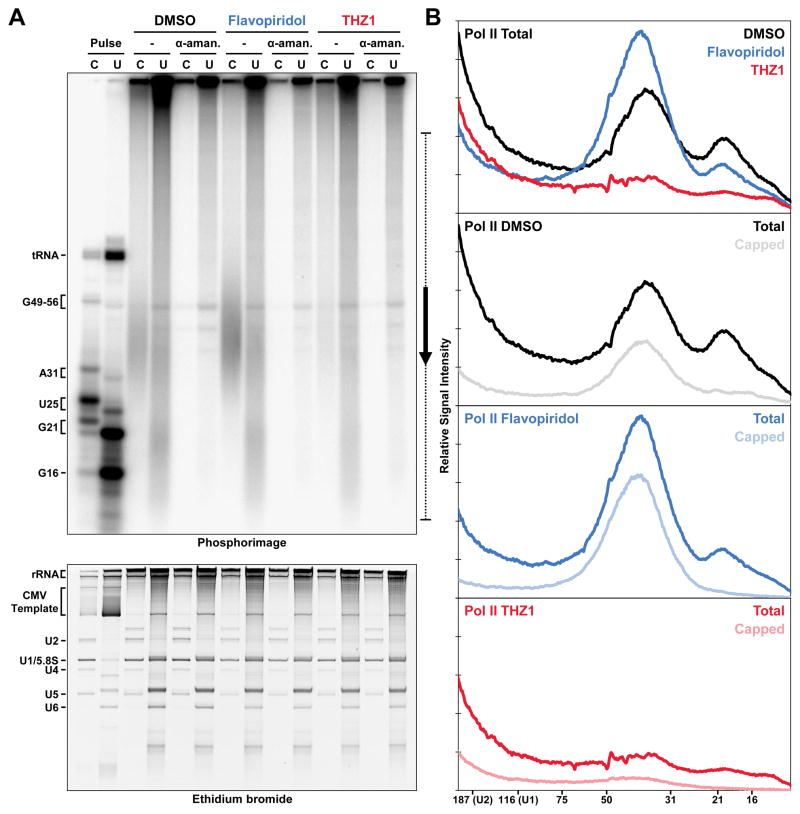
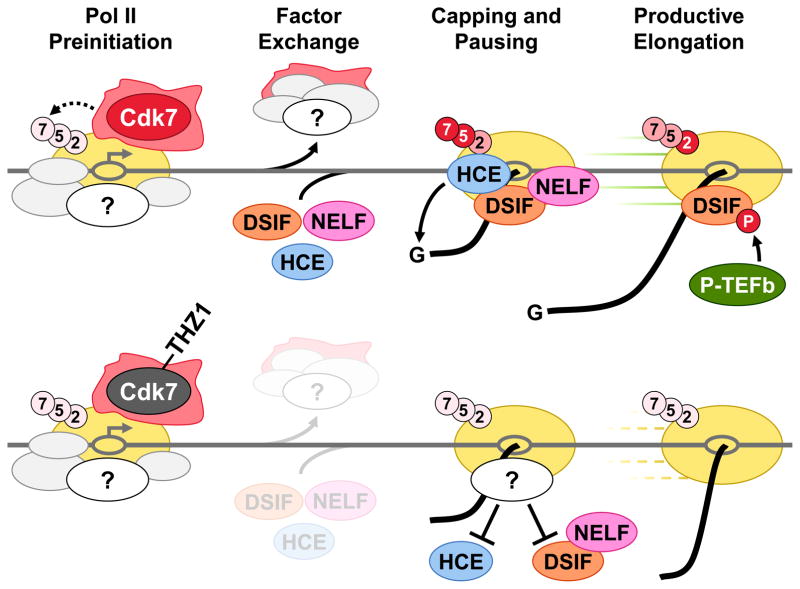
The Cdk7 subunit of TFIIH phosphorylates RNA polymerase II (Pol II) during initiation, and, while recent studies show that inhibition of human Cdk7 negatively influences transcription, the mechanisms involved are unclear. Using in vitro transcription with nuclear extract, we demonstrate that THZ1, a covalent Cdk7 inhibitor, causes defects in Pol II phosphorylation, co-transcriptional capping, promoter proximal pausing, and productive elongation. THZ1 does not affect initiation but blocks essentially all Pol II large subunit C-terminal domain (CTD) phosphorylation. We found that guanylylation of nascent RNAs is length dependent and modulated by a THZ1-sensitive factor present in nuclear extract. THZ1 impacts pausing through a capping-independent block of DSIF and NELF loading. The P-TEFb-dependent transition into productive elongation was also inhibited by THZ1, likely due to loss of DSIF. Capping and pausing were also reduced in THZ1-treated cells. Our results provide mechanistic insights into THZ1 action and how Cdk7 broadly influences transcription and capping.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Revisiting the Function of CDK7 in Transcription by Virtue of a Recently Described TFIIH Kinase Inhibitor.Mol Cell. 2015 Aug 20;59(4):513-4. doi: 10.1016/j.molcel.2015.08.006. Mol Cell. 2015. PMID: 26295956
References
-
- Adamson TE, Shore SM, Price DH. Analysis of RNA polymerase II elongation in vitro. Methods Enzymol. 2003;371:264–275. - PubMed
-
- Akoulitchev S, Makela TP, Weinberg RA, Reinberg D. Requirement for TFIIH kinase activity in transcription by RNA polymerase II. Nature. 1995;377:557–560. - PubMed
-
- Bataille AR, Jeronimo C, Jacques PE, Laramee L, Fortin ME, Forest A, Bergeron M, Hanes SD, Robert F. A universal RNA polymerase II CTD cycle is orchestrated by complex interplays between kinase, phosphatase, and isomerase enzymes along genes. Mol Cell. 2012;45:158–170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
