

Investigating neuronal function with optically controllable proteins
- PMID: 26257603
- PMCID: PMC4508517
- DOI: 10.3389/fnmol.2015.00037
Investigating neuronal function with optically controllable proteins
Abstract
In the nervous system, protein activities are highly regulated in space and time. This regulation allows for fine modulation of neuronal structure and function during development and adaptive responses. For example, neurite extension and synaptogenesis both involve localized and transient activation of cytoskeletal and signaling proteins, allowing changes in microarchitecture to occur rapidly and in a localized manner. To investigate the role of specific protein regulation events in these processes, methods to optically control the activity of specific proteins have been developed. In this review, we focus on how photosensory domains enable optical control over protein activity and have been used in neuroscience applications. These tools have demonstrated versatility in controlling various proteins and thereby cellular functions, and possess enormous potential for future applications in nervous systems. Just as optogenetic control of neuronal firing using opsins has changed how we investigate the function of cellular circuits in vivo, optical control may yet yield another revolution in how we study the circuitry of intracellular signaling in the brain.
Keywords: development; optobiology; optogenetics; signal transduction; transcription.
Figures

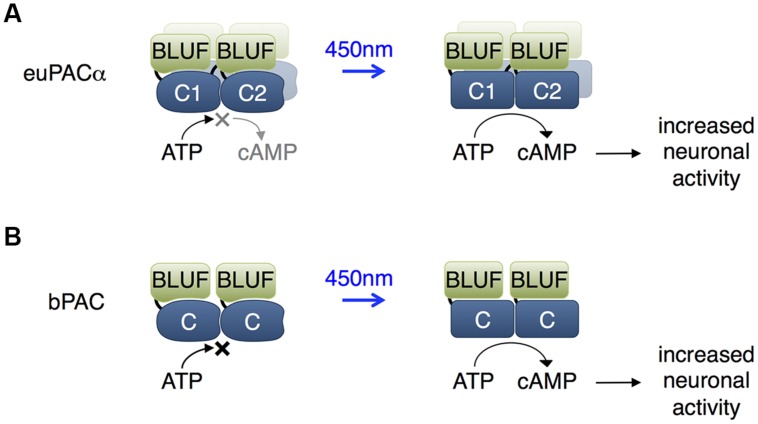



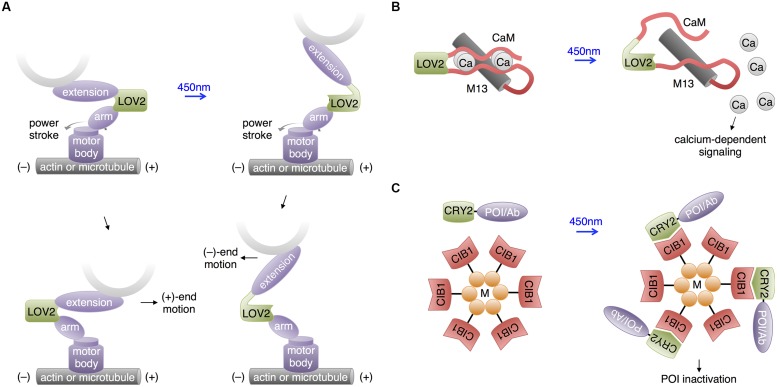

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources