

Genetic control of inflorescence architecture in legumes
- PMID: 26257753
- PMCID: PMC4508509
- DOI: 10.3389/fpls.2015.00543
Genetic control of inflorescence architecture in legumes
Abstract
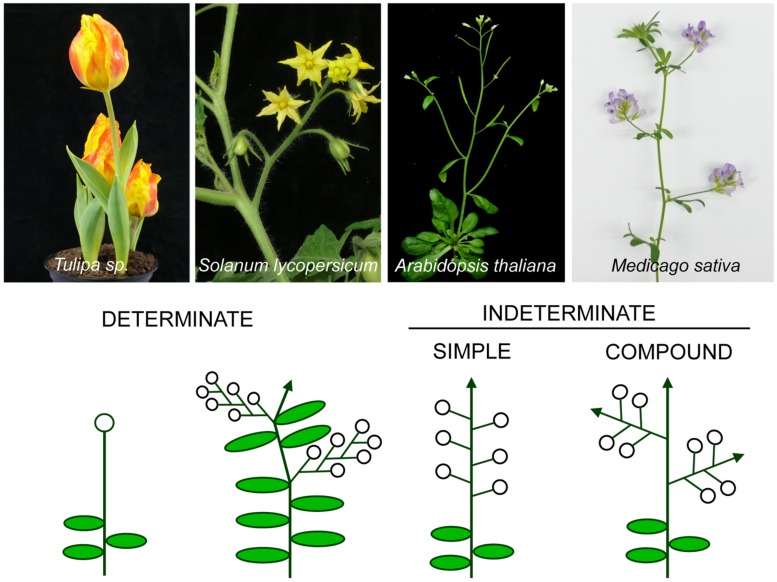
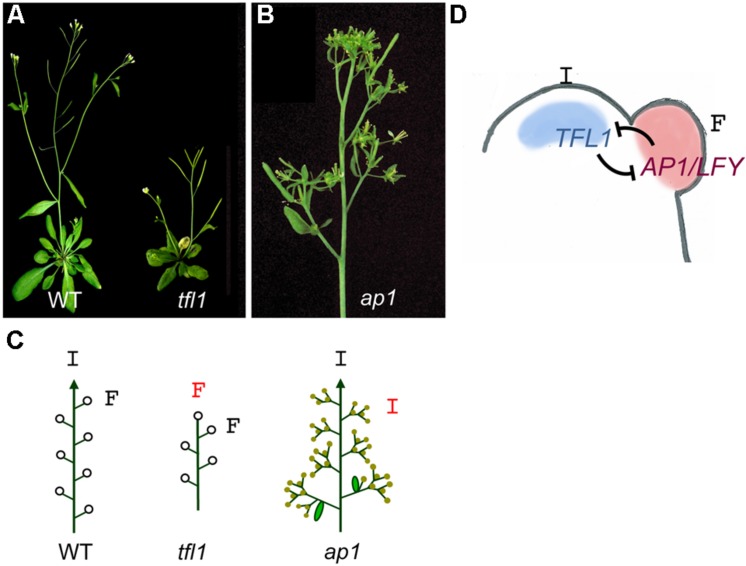
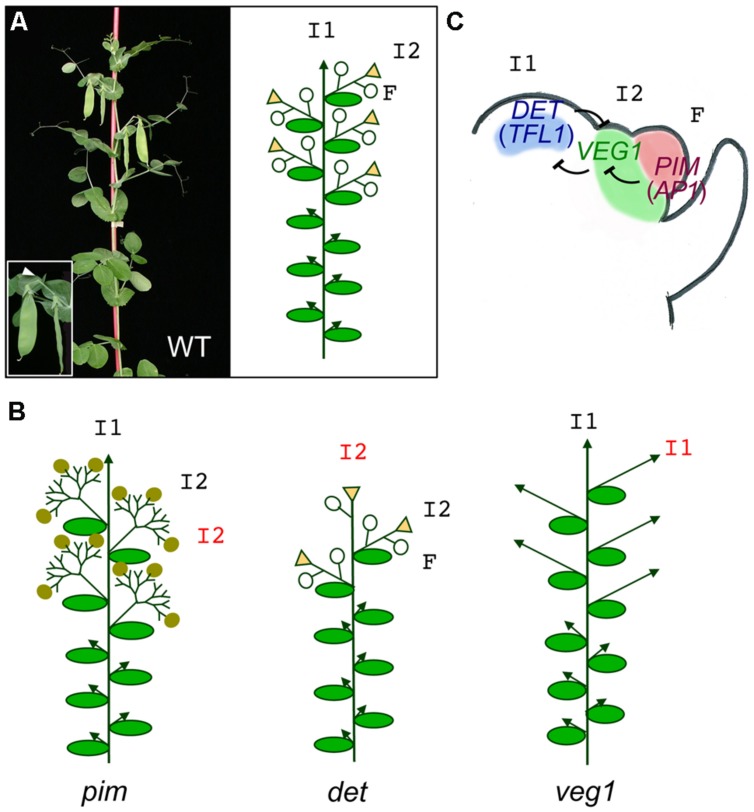
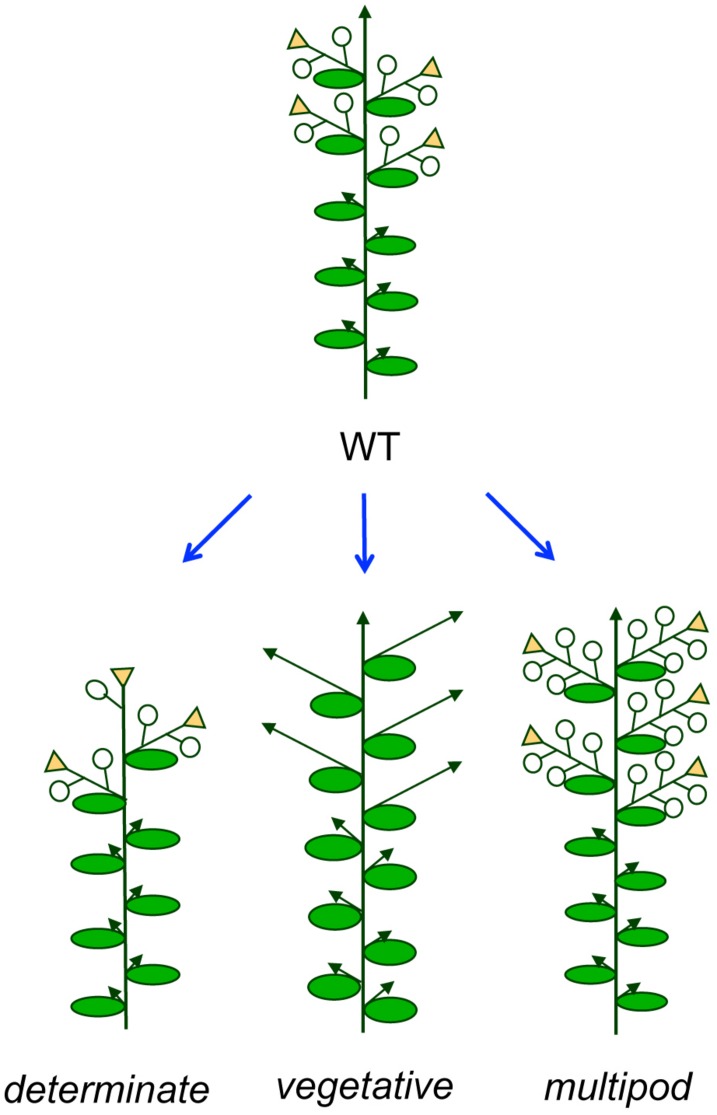
The architecture of the inflorescence, the shoot system that bears the flowers, is a main component of the huge diversity of forms found in flowering plants. Inflorescence architecture has also a strong impact on the production of fruits and seeds, and on crop management, two highly relevant agronomical traits. Elucidating the genetic networks that control inflorescence development, and how they vary between different species, is essential to understanding the evolution of plant form and to being able to breed key architectural traits in crop species. Inflorescence architecture depends on the identity and activity of the meristems in the inflorescence apex, which determines when flowers are formed, how many are produced and their relative position in the inflorescence axis. Arabidopsis thaliana, where the genetic control of inflorescence development is best known, has a simple inflorescence, where the primary inflorescence meristem directly produces the flowers, which are thus borne in the main inflorescence axis. In contrast, legumes represent a more complex inflorescence type, the compound inflorescence, where flowers are not directly borne in the main inflorescence axis but, instead, they are formed by secondary or higher order inflorescence meristems. Studies in model legumes such as pea (Pisum sativum) or Medicago truncatula have led to a rather good knowledge of the genetic control of the development of the legume compound inflorescence. In addition, the increasing availability of genetic and genomic tools for legumes is allowing to rapidly extending this knowledge to other grain legume crops. This review aims to describe the current knowledge of the genetic network controlling inflorescence development in legumes. It also discusses how the combination of this knowledge with the use of emerging genomic tools and resources may allow rapid advances in the breeding of grain legume crops.
Keywords: AP1; TFL1; VEG1; inflorescence architecture; legumes; meristem identity; pea.
Figures
References
-
- Alvarez J., Guli C. L., Yu X. H., Smyth D. R. (1992). terminal flower: a gene affecting inflorescence development in Arabidopsis thaliana. Plant J. 2 103–116. 10.1111/j.1365-313X.1992.00103.x - DOI
-
- Annicchiarico P., Iannucci A. (2008). Adaptation strategy, germplasm type and adaptive traits for field pea improvement in Italy based on variety responses across climatically contrasting environments. Field Crops Res. 108 133–142. 10.1016/j.fcr.2008.04.004 - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous