

Involvement of adiponectin in the pathogenesis of dystrophinopathy
- PMID: 26257862
- PMCID: PMC4528853
- DOI: 10.1186/s13395-015-0051-9
Involvement of adiponectin in the pathogenesis of dystrophinopathy
Abstract
Background: The hormone adiponectin (ApN) is decreased in the metabolic syndrome, where it plays a key pathogenic role. ApN also exerts some anti-inflammatory effects on skeletal muscles in mice exposed to acute or chronic inflammation. Here, we investigate whether ApN could be sufficiently potent to counteract a severe degenerative muscle disease, with an inflammatory component such as Duchenne muscular dystrophy (DMD).
Methods: Mdx mice (a DMD model caused by dystrophin mutation) were crossed with mice overexpressing ApN in order to generate mdx-ApN mice; only littermates were used. Different markers of inflammation/oxidative stress and components of signaling pathways were studied. Global force was assessed by in vivo functional tests, and muscle injury with Evans Blue Dye (EBD). Eventually, primary cultures of human myotubes were used.
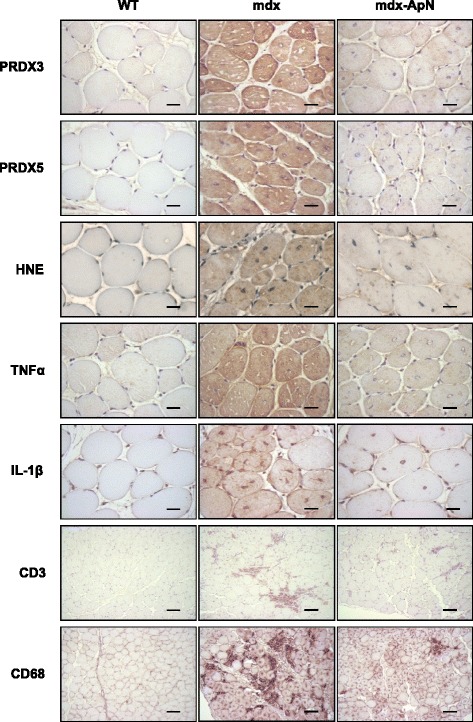
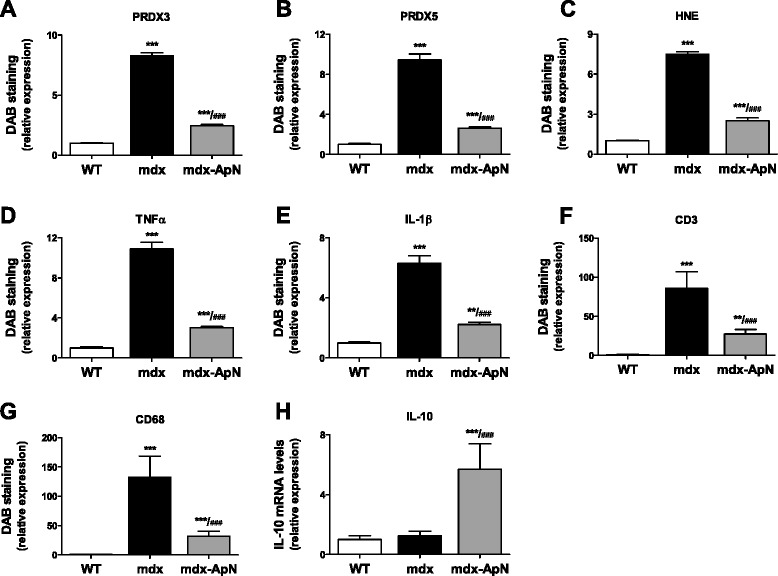
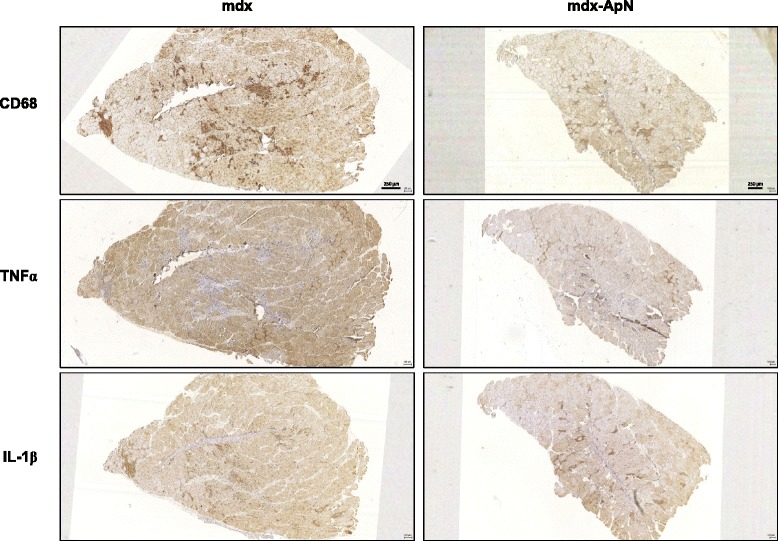
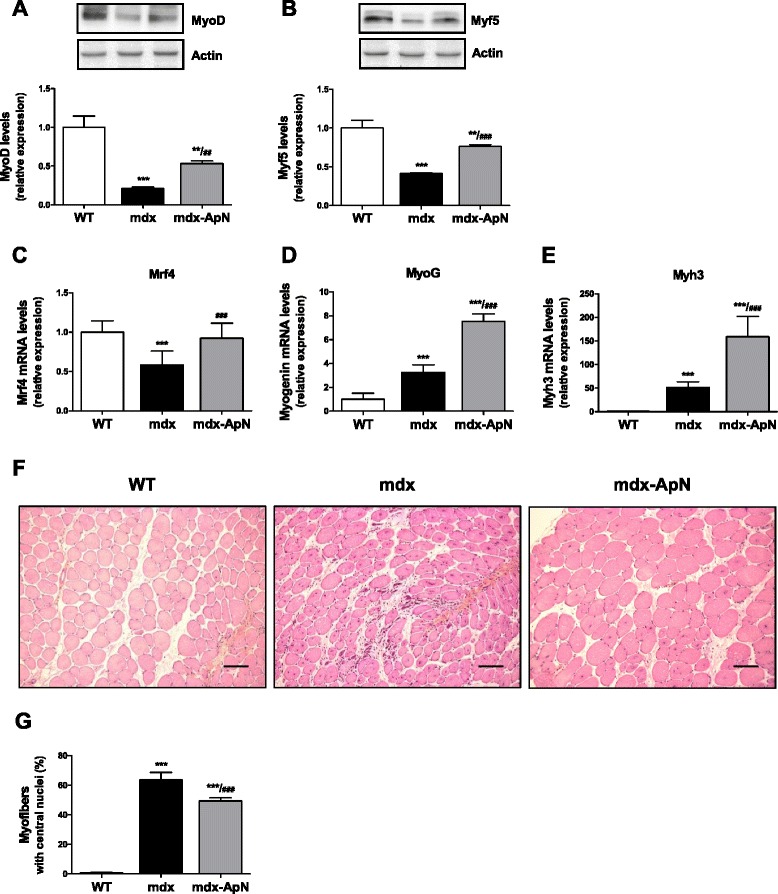
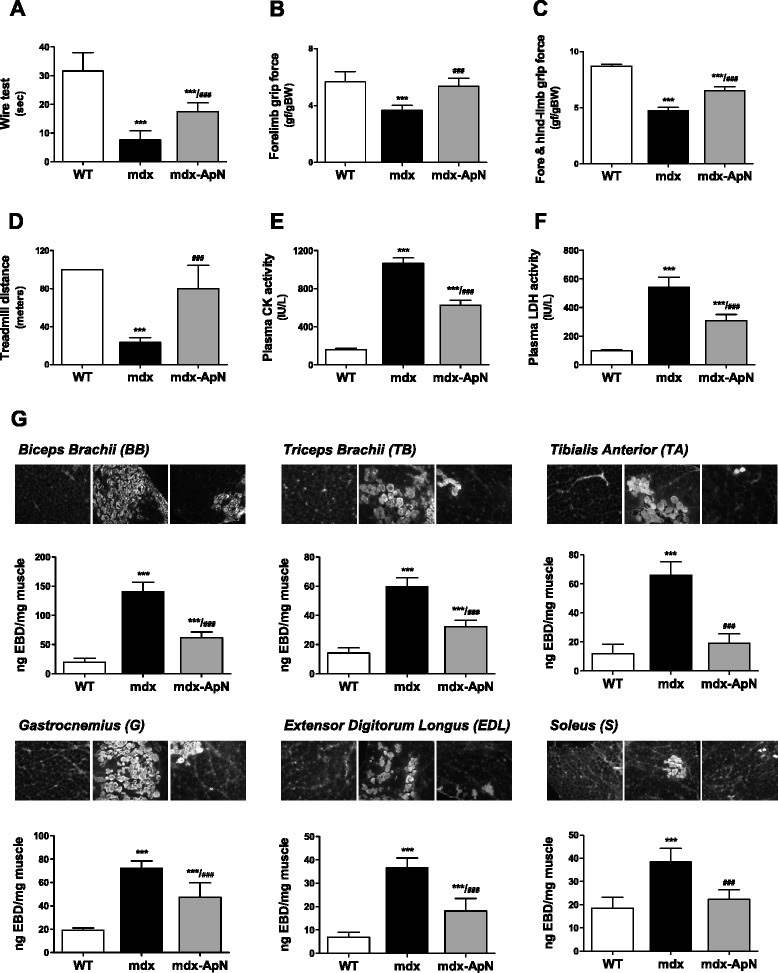
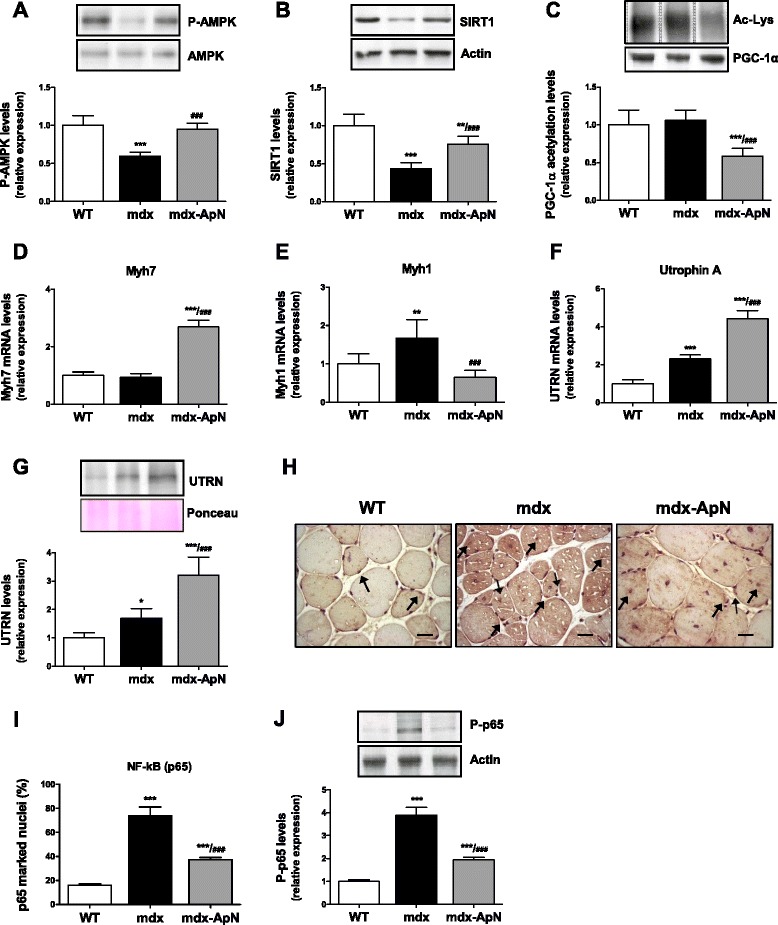
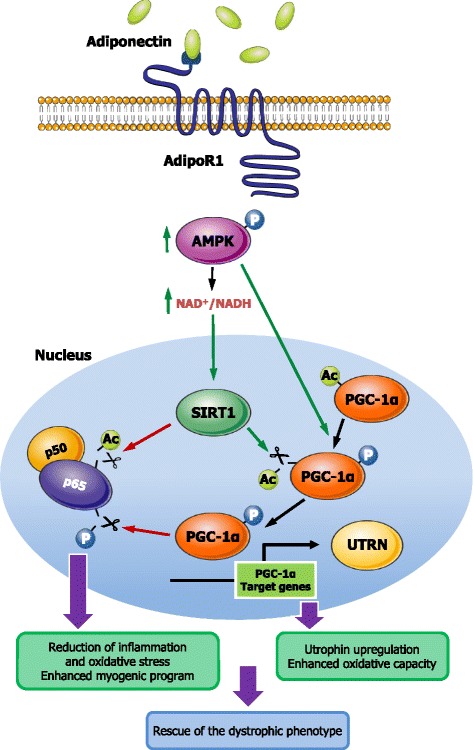
Results: Circulating ApN was markedly diminished in mdx mice. Replenishment of ApN strikingly reduced muscle inflammation, oxidative stress, and enhanced the expression of myogenic differentiation markers along with that of utrophin A (a dystrophin analog) in mdx-ApN mice. Accordingly, mdx-ApN mice exhibited higher global force and endurance as well as decreased muscle damage as quantified by curtailed extravasation of EBD in myofibers. These beneficial effects of ApN were recapitulated in human myotubes. ApN mediates its protection via the adiponectin receptor 1 (AdipoR1, the main ApN receptor in muscle) and the AMPK-SIRT1-PGC-1α signaling pathway, leading to downregulation of the nuclear factor kappa B (NF-κB) and inflammatory genes, together with upregulation of utrophin.
Conclusions: Adiponectin proves to be an extremely powerful hormone capable of protecting the skeletal muscle against inflammation and injury, thereby offering novel therapeutic perspectives for dystrophinopathies.
Keywords: AMPK signaling; Adiponectin; Inflammation; Mdx; NF-κB; Skeletal muscle; Utrophin.
Figures

References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
