

Mutations in DCHS1 cause mitral valve prolapse
- PMID: 26258302
- PMCID: PMC4720389
- DOI: 10.1038/nature14670
Mutations in DCHS1 cause mitral valve prolapse
Abstract
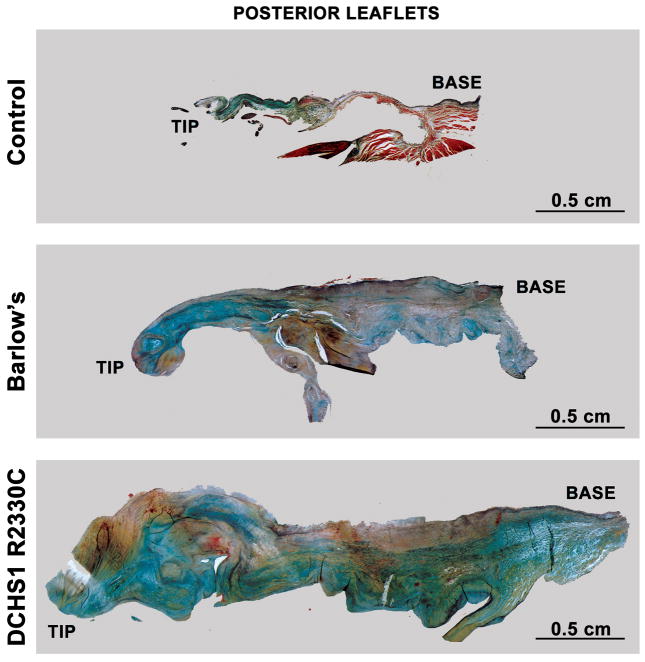
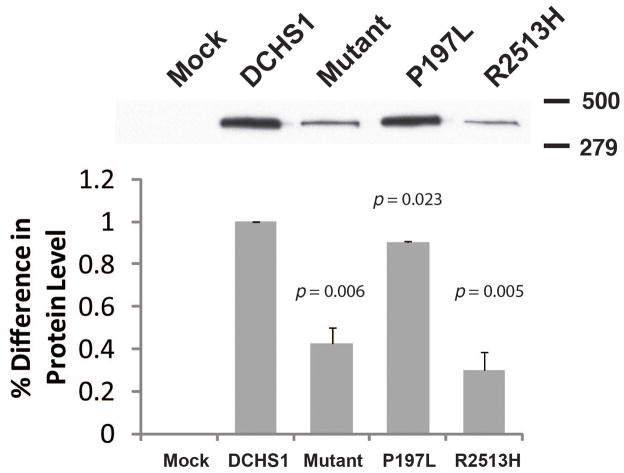
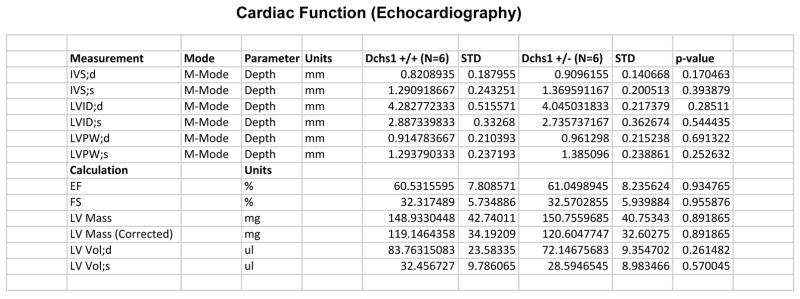
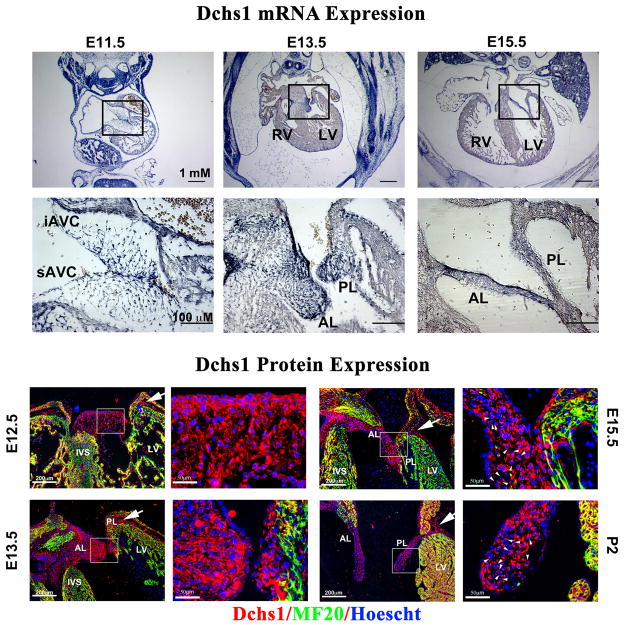
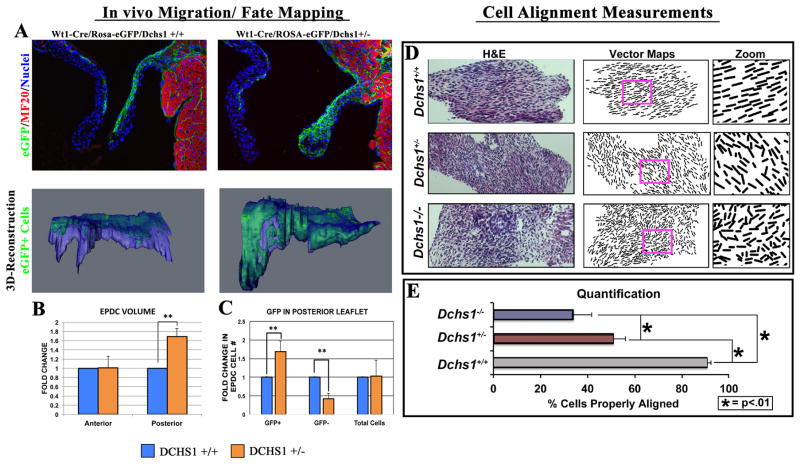
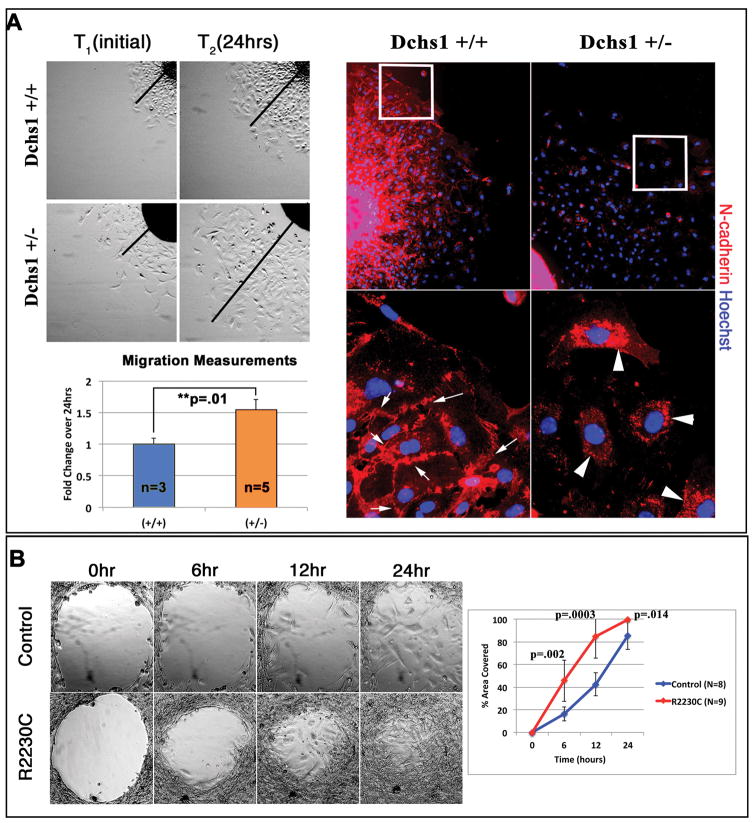
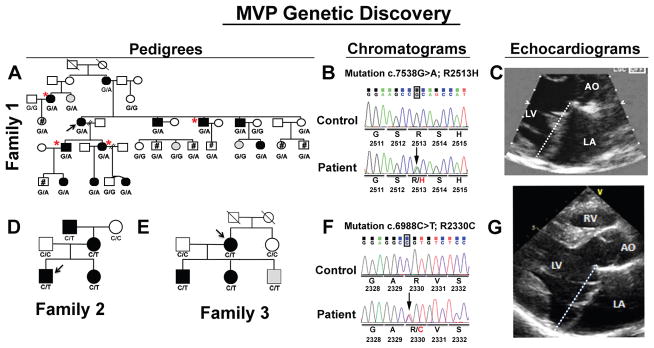
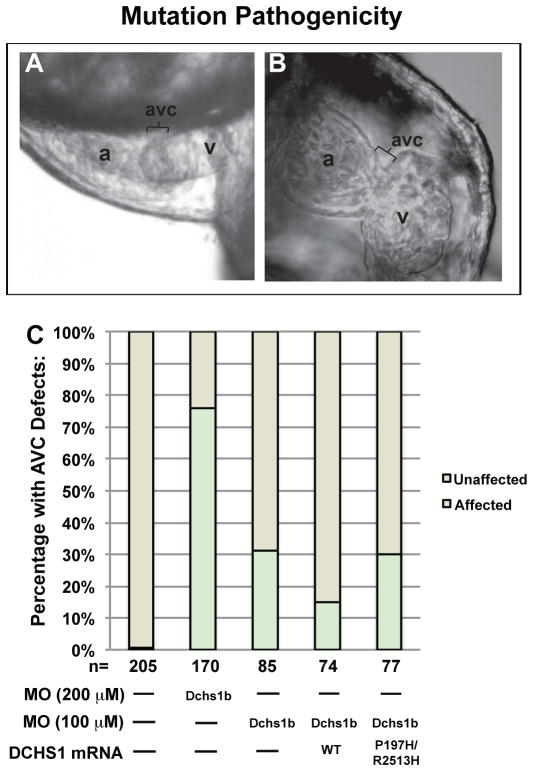
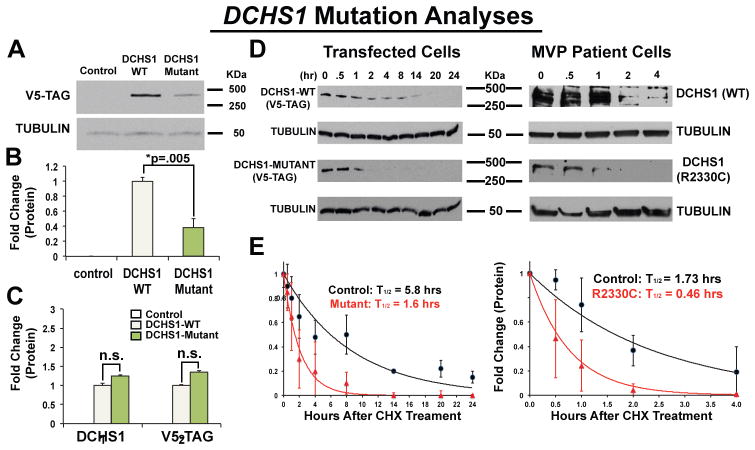
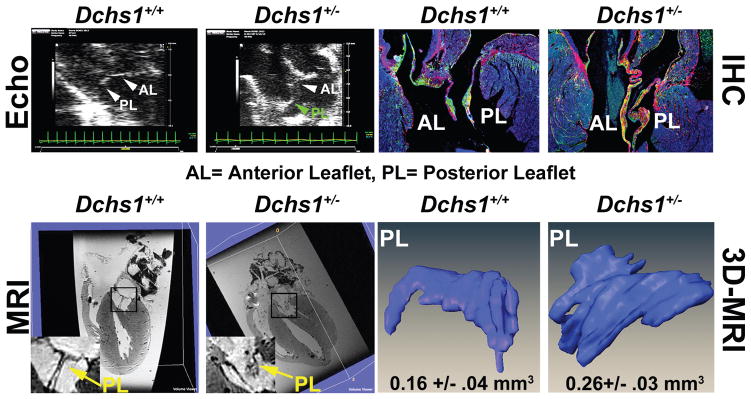
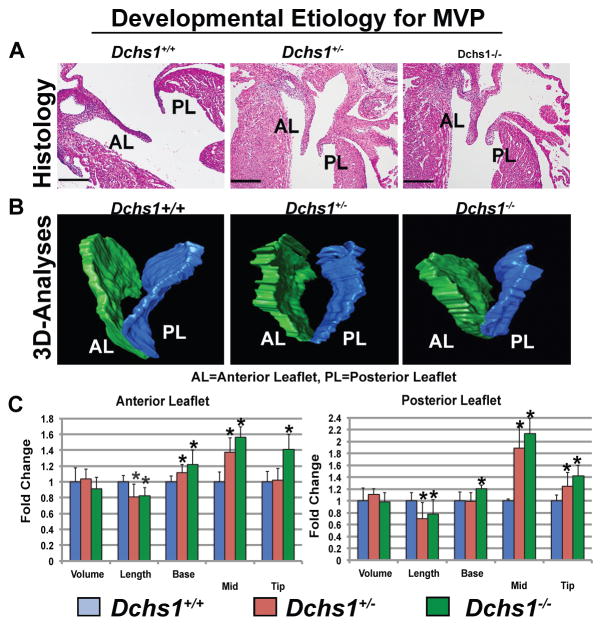
Mitral valve prolapse (MVP) is a common cardiac valve disease that affects nearly 1 in 40 individuals. It can manifest as mitral regurgitation and is the leading indication for mitral valve surgery. Despite a clear heritable component, the genetic aetiology leading to non-syndromic MVP has remained elusive. Four affected individuals from a large multigenerational family segregating non-syndromic MVP underwent capture sequencing of the linked interval on chromosome 11. We report a missense mutation in the DCHS1 gene, the human homologue of the Drosophila cell polarity gene dachsous (ds), that segregates with MVP in the family. Morpholino knockdown of the zebrafish homologue dachsous1b resulted in a cardiac atrioventricular canal defect that could be rescued by wild-type human DCHS1, but not by DCHS1 messenger RNA with the familial mutation. Further genetic studies identified two additional families in which a second deleterious DCHS1 mutation segregates with MVP. Both DCHS1 mutations reduce protein stability as demonstrated in zebrafish, cultured cells and, notably, in mitral valve interstitial cells (MVICs) obtained during mitral valve repair surgery of a proband. Dchs1(+/-) mice had prolapse of thickened mitral leaflets, which could be traced back to developmental errors in valve morphogenesis. DCHS1 deficiency in MVP patient MVICs, as well as in Dchs1(+/-) mouse MVICs, result in altered migration and cellular patterning, supporting these processes as aetiological underpinnings for the disease. Understanding the role of DCHS1 in mitral valve development and MVP pathogenesis holds potential for therapeutic insights for this very common disease.
Conflict of interest statement
Figures
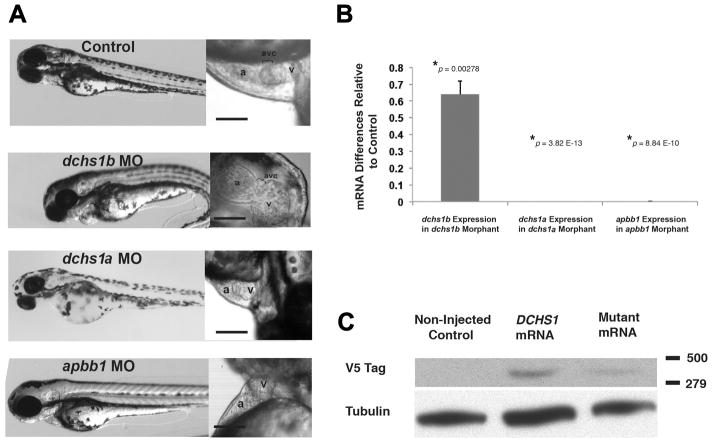
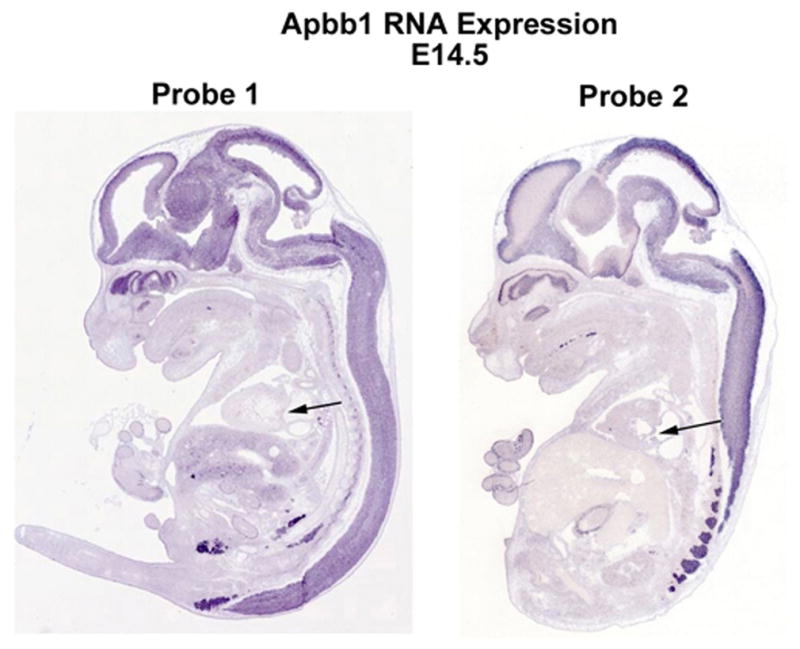
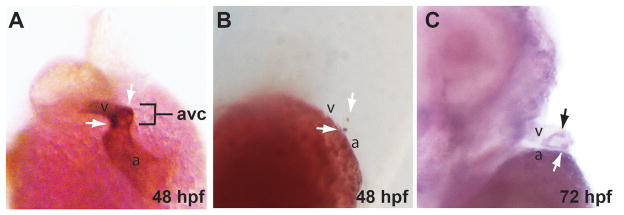
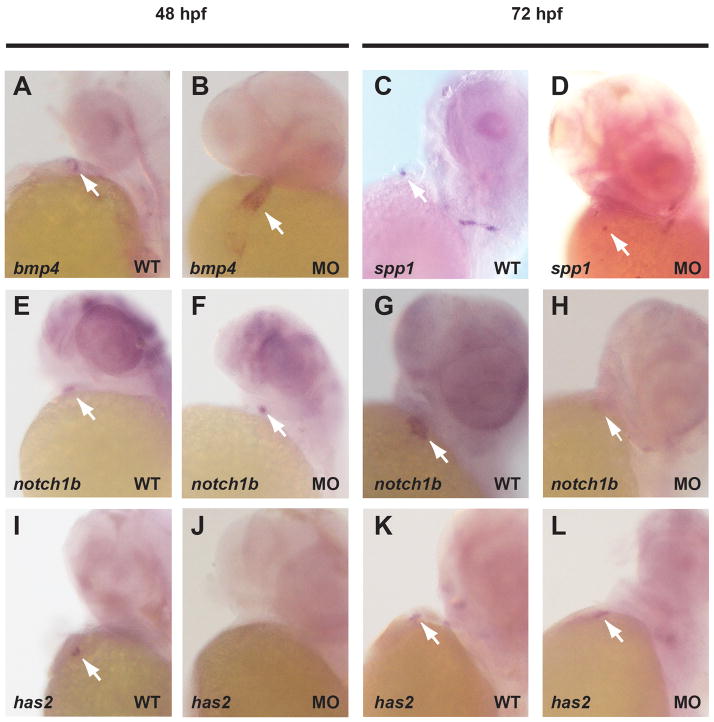
References
-
- Freed LA, et al. Mitral valve prolapse in the general population: the benign nature of echocardiographic features in the Framingham Heart Study. J Am Coll Cardiol. 2002;40:1298–1304. S0735109702021617 [pii] - PubMed
-
- Avierinos JF, et al. Natural history of asymptomatic mitral valve prolapse in the community. Circulation. 2002;106:1355–1361. - PubMed
-
- Waller BF, Maron BJ, Del Negro AA, Gottdiener JS, Roberts WC. Frequency and significance of M-mode echocardiographic evidence of mitral valve prolapse in clinically isolated pure mitral regurgitation: analysis of 65 patients having mitral valve replacement. The American journal of cardiology. 1984;53:139–147. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- K24 HL067434/HL/NHLBI NIH HHS/United States
- R01 HL109506/HL/NHLBI NIH HHS/United States
- R01-HL127692/HL/NHLBI NIH HHS/United States
- R01 HL127692/HL/NHLBI NIH HHS/United States
- R01HL122906-01/HL/NHLBI NIH HHS/United States
- R00 MH095867/MH/NIMH NIH HHS/United States
- R01 HL122906/HL/NHLBI NIH HHS/United States
- P20 GM103444/GM/NIGMS NIH HHS/United States
- 8P20 GM103444-07/GM/NIGMS NIH HHS/United States
- K24 HL67434/HL/NHLBI NIH HHS/United States
- R01 HL128099/HL/NHLBI NIH HHS/United States
- K99 MH095867/MH/NIMH NIH HHS/United States
- R01 HL033756/HL/NHLBI NIH HHS/United States
- R01HL109506/HL/NHLBI NIH HHS/United States
- T32 HL007208/HL/NHLBI NIH HHS/United States
- P30 GM103342/GM/NIGMS NIH HHS/United States
- R01 HL072265/HL/NHLBI NIH HHS/United States
- C06 RR018823/RR/NCRR NIH HHS/United States
- R00-MH095867/MH/NIMH NIH HHS/United States
- R01 HL109004/HL/NHLBI NIH HHS/United States
- R01-HL095696/HL/NHLBI NIH HHS/United States
- Howard Hughes Medical Institute/United States
- R01HL72265/HL/NHLBI NIH HHS/United States
- R01-HL109004/HL/NHLBI NIH HHS/United States
- 1P30 GM103342/GM/NIGMS NIH HHS/United States
- I01 BX002327/BX/BLRD VA/United States
- K23 HL116652/HL/NHLBI NIH HHS/United States
- R01-HL33756/HL/NHLBI NIH HHS/United States
- T32 HL007260/HL/NHLBI NIH HHS/United States
- R01 HL095696/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
