

An excess of catalytically required motions inhibits the scavenger decapping enzyme
- PMID: 26258763
- PMCID: PMC4544744
- DOI: 10.1038/nchembio.1866
An excess of catalytically required motions inhibits the scavenger decapping enzyme
Abstract
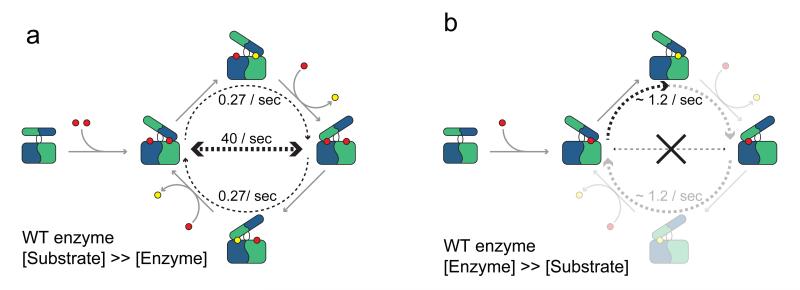
The scavenger decapping enzyme hydrolyzes the protective 5' cap structure on short mRNA fragments that are generated from the exosomal degradation of mRNAs. From static crystal structures and NMR data, it is apparent that the dimeric enzyme has to undergo large structural changes to bind its substrate in a catalytically competent conformation. Here we studied the yeast enzyme and showed that the associated opening and closing motions can be orders of magnitude faster than the catalytic turnover rate. This excess of motion is induced by the binding of a second ligand to the enzyme, which occurs at high substrate concentrations. We designed a mutant that disrupted the allosteric pathway that links the second binding event to the dynamics and showed that this mutant enzyme is hyperactive. Our data reveal a unique mechanism of substrate inhibition in which motions that are required for catalytic activity also inhibit efficient turnover when they are present in excess.
Figures




References
-
- Fersht AR. Structure and mechanism in protein science: A guide to enzyme catalysis and protein folding. W. H. Freeman; 1999.
-
- Hammes-Schiffer S, Benkovic SJ. Relating protein motion to catalysis. Annu Rev Biochem. 2006;75:519–41. - PubMed
-
- Boehr DD, Dyson HJ, Wright PE. An NMR perspective on enzyme dynamics. Chem Rev. 2006;106:3055–79. - PubMed
-
- Henzler-Wildman K, Kern D. Dynamic personalities of proteins. Nature. 2007;450:964–72. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
