

β-catenin stabilization enhances SS18-SSX2-driven synovial sarcomagenesis and blocks the mesenchymal to epithelial transition
- PMID: 26259251
- PMCID: PMC4673197
- DOI: 10.18632/oncotarget.4283
β-catenin stabilization enhances SS18-SSX2-driven synovial sarcomagenesis and blocks the mesenchymal to epithelial transition
Abstract
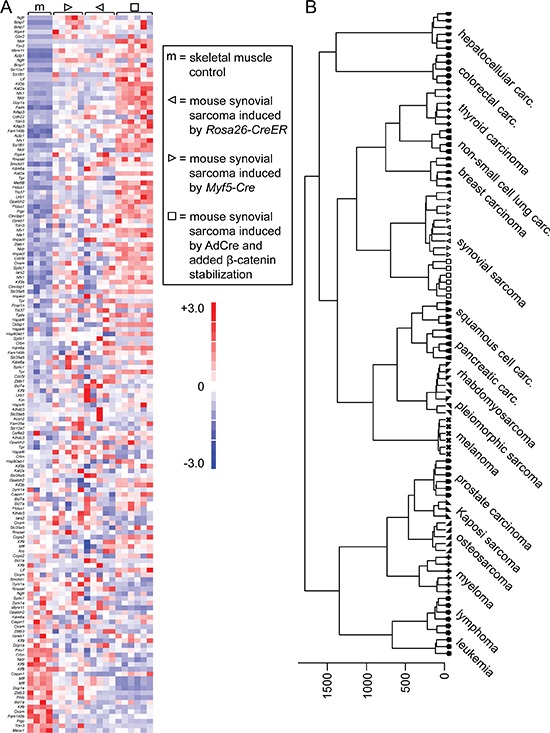
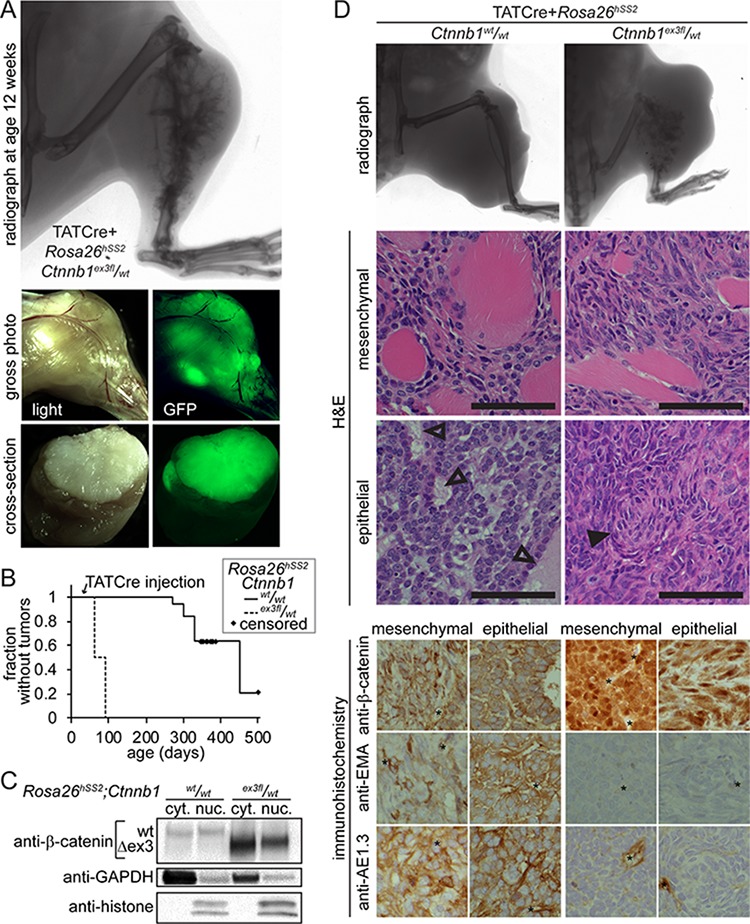
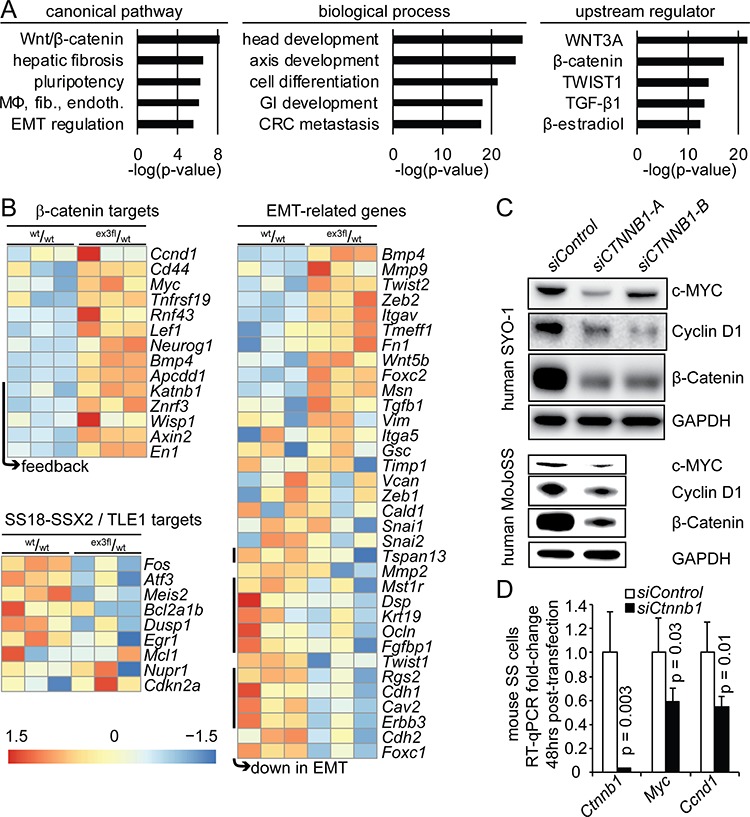
β-catenin is a master regulator in the cellular biology of development and neoplasia. Its dysregulation is implicated as a driver of colorectal carcinogenesis and the epithelial-mesenchymal transition in other cancers. Nuclear β-catenin staining is a poor prognostic sign in synovial sarcoma, the most common soft-tissue sarcoma in adolescents and young adults. We show through genetic experiments in a mouse model that expression of a stabilized form of β-catenin greatly enhances synovial sarcomagenesis. Stabilization of β-catenin enables a stem-cell phenotype in synovial sarcoma cells, specifically blocking epithelial differentiation and driving invasion. β-catenin achieves its reprogramming in part by upregulating transcription of TCF/LEF target genes. Even though synovial sarcoma is primarily a mesenchymal neoplasm, its progression towards a more aggressive and invasive phenotype parallels the epithelial-mesenchymal transition observed in epithelial cancers, where β-catenin's transcriptional contribution includes blocking epithelial differentiation.
Keywords: Wnt-signaling; epithelial-mesenchymal transition; mouse genetic model; translocation.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures

References
-
- Herzog CE. Overview of sarcomas in the adolescent and young adult population. J Pediatr Hematol Oncol. 2005;27:215–8. - PubMed
-
- Ladanyi M, Antonescu CR, Leung DH, Woodruff JM, Kawai A, Healey JH, Brennan MF, Bridge JA, Neff JR, Barr FG, Goldsmith JD, Brooks JS, Goldblum JR, Ali SZ, Shipley J, Cooper CS, Fisher C, Skytting B, Larsson O. Impact of SYT-SSX fusion type on the clinical behavior of synovial sarcoma: a multi-institutional retrospective study of 243 patients. Cancer Res. 2002;62:135–40. - PubMed
-
- Haldar M, Hancock JD, Coffin CM, Lessnick SL, Capecchi MR. A conditional mouse model of synovial sarcoma: insights into a myogenic origin. Cancer cell. 2007;11:375–88. - PubMed
-
- Moon RT, Kohn AD, De Ferrari GV, Kaykas A. WNT and beta-catenin signalling: diseases and therapies. Nat Rev Genet. 2004;5:691–701. - PubMed
-
- Liu C, Li Y, Semenov M, Han C, Baeg GH, Tan Y, Zhang Z, Lin X, He X. Control of beta-catenin phosphorylation/degradation by a dual-kinase mechanism. Cell. 2002;108:837–47. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
