

Initiation of mRNA translation in bacteria: structural and dynamic aspects
- PMID: 26259514
- PMCID: PMC4611024
- DOI: 10.1007/s00018-015-2010-3
Initiation of mRNA translation in bacteria: structural and dynamic aspects
Abstract
Initiation of mRNA translation is a major checkpoint for regulating level and fidelity of protein synthesis. Being rate limiting in protein synthesis, translation initiation also represents the target of many post-transcriptional mechanisms regulating gene expression. The process begins with the formation of an unstable 30S pre-initiation complex (30S pre-IC) containing initiation factors (IFs) IF1, IF2 and IF3, the translation initiation region of an mRNA and initiator fMet-tRNA whose codon and anticodon pair in the P-site following a first-order rearrangement of the 30S pre-IC produces a locked 30S initiation complex (30SIC); this is docked by the 50S subunit to form a 70S complex that, following several conformational changes, positional readjustments of its ligands and ejection of the IFs, becomes a 70S initiation complex productive in initiation dipeptide formation. The first EF-G-dependent translocation marks the beginning of the elongation phase of translation. Here, we review structural, mechanistic and dynamical aspects of this process.
Keywords: GTP; Protein synthesis; Translation initiation factors; fMet-tRNA; mRNA initiation region.
Figures



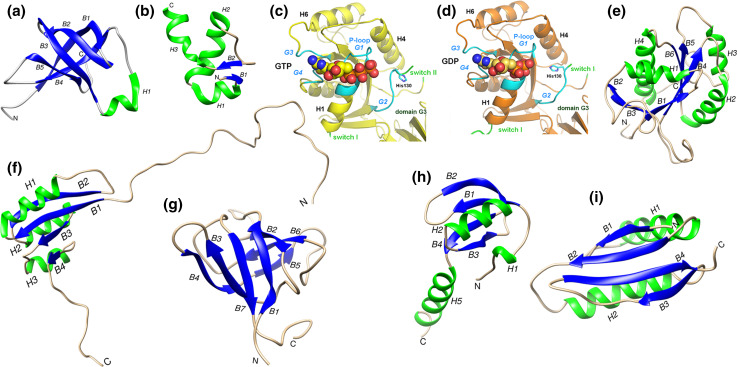

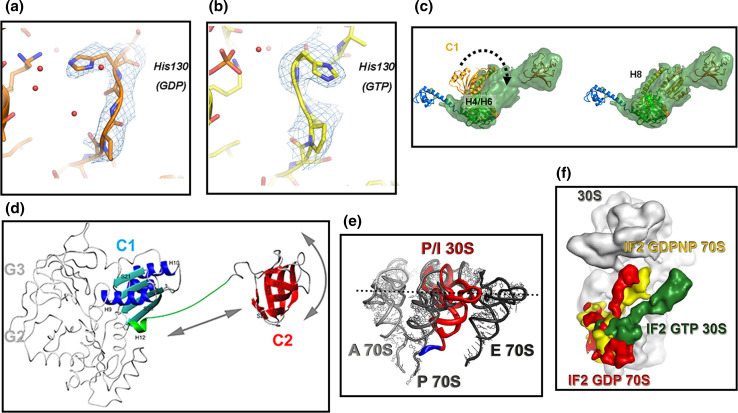

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
