

Reciprocal regulation of two G protein-coupled receptors sensing extracellular concentrations of Ca2+ and H
- PMID: 26261299
- PMCID: PMC4553783
- DOI: 10.1073/pnas.1506085112
Reciprocal regulation of two G protein-coupled receptors sensing extracellular concentrations of Ca2+ and H
Abstract
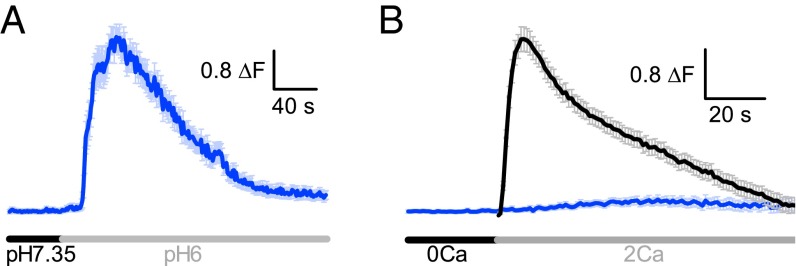
G protein-coupled receptors (GPCRs) are cell surface receptors that detect a wide range of extracellular messengers and convey this information to the inside of cells. Extracellular calcium-sensing receptor (CaSR) and ovarian cancer gene receptor 1 (OGR1) are two GPCRs that sense extracellular Ca(2+) and H(+), respectively. These two ions are key components of the interstitial fluid, and their concentrations change in an activity-dependent manner. Importantly, the interstitial fluid forms part of the microenvironment that influences cell function in health and disease; however, the exact mechanisms through which changes in the microenvironment influence cell function remain largely unknown. We show that CaSR and OGR1 reciprocally inhibit signaling through each other in central neurons, and that this is lost in their transformed counterparts. Furthermore, strong intracellular acidification impairs CaSR function, but potentiates OGR1 function. Thus, CaSR and OGR1 activities can be regulated in a seesaw manner, whereby conditions promoting signaling through one receptor simultaneously inhibit signaling through the other receptor, potentiating the difference in their relative signaling activity. Our results provide insight into how small but consistent changes in the ionic microenvironment of cells can significantly alter the balance between two signaling pathways, which may contribute to disease progression.
Keywords: CaSR; OGR1; extracellular acidosis; microenvironment; pH sensing.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
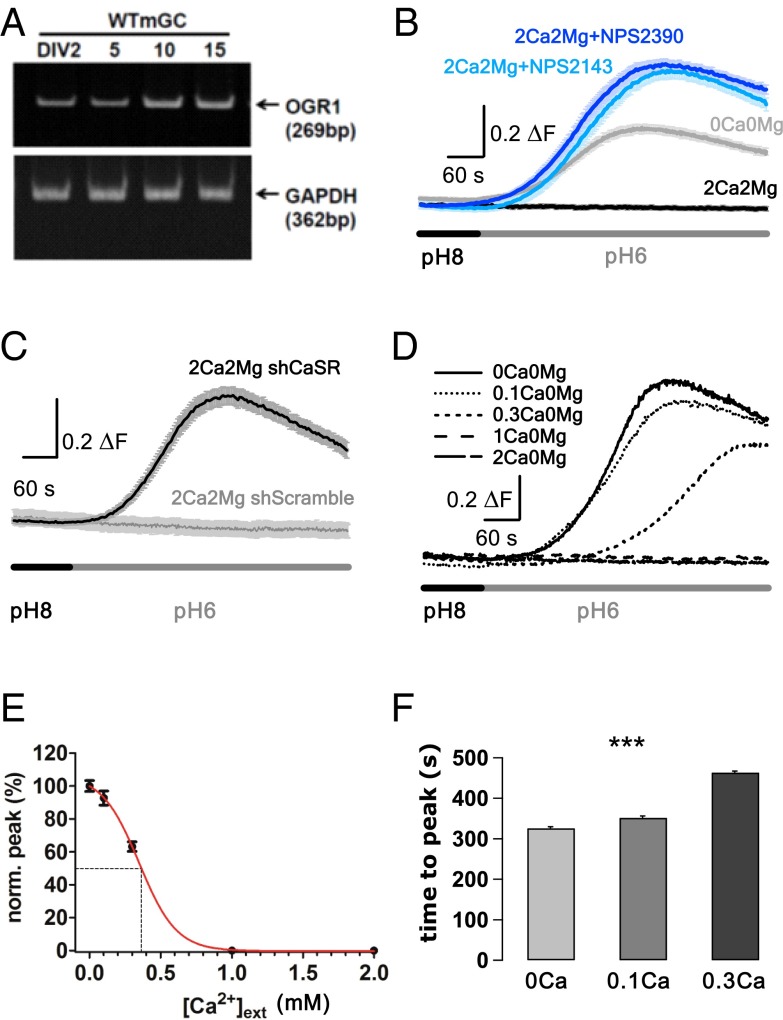
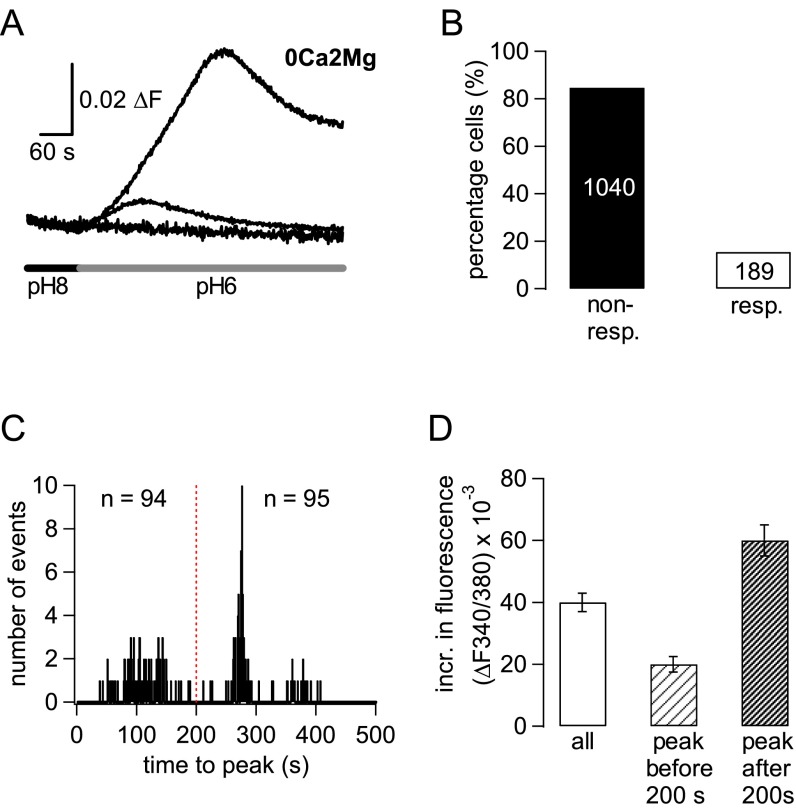
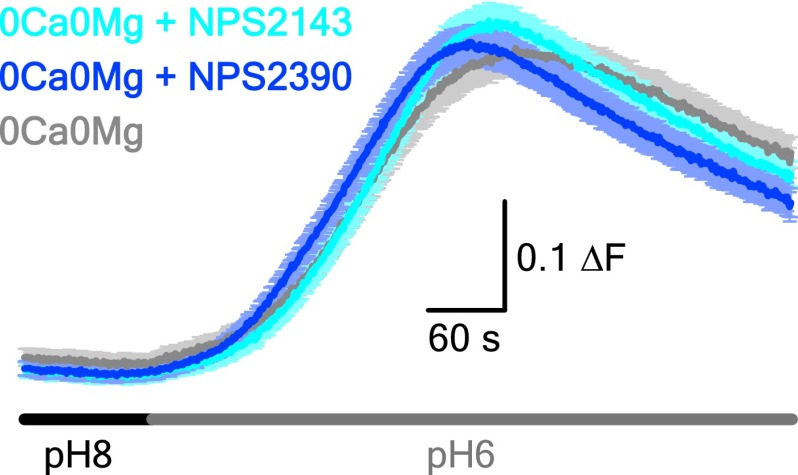
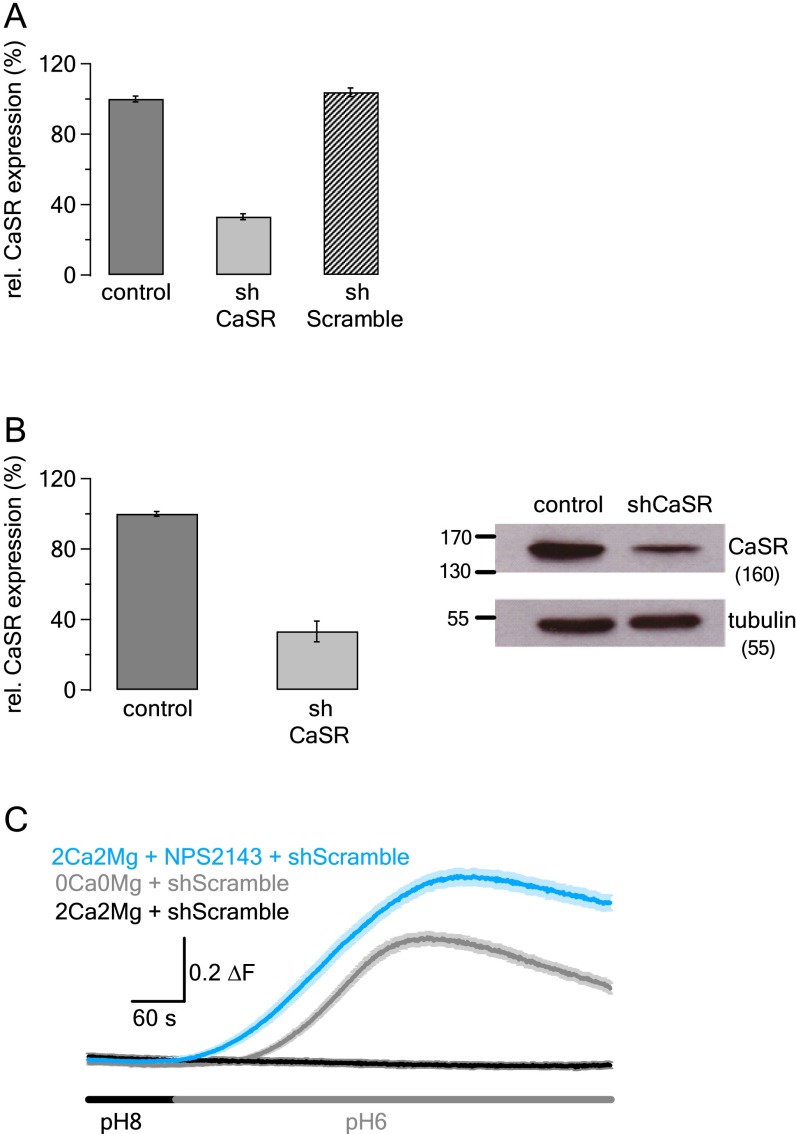
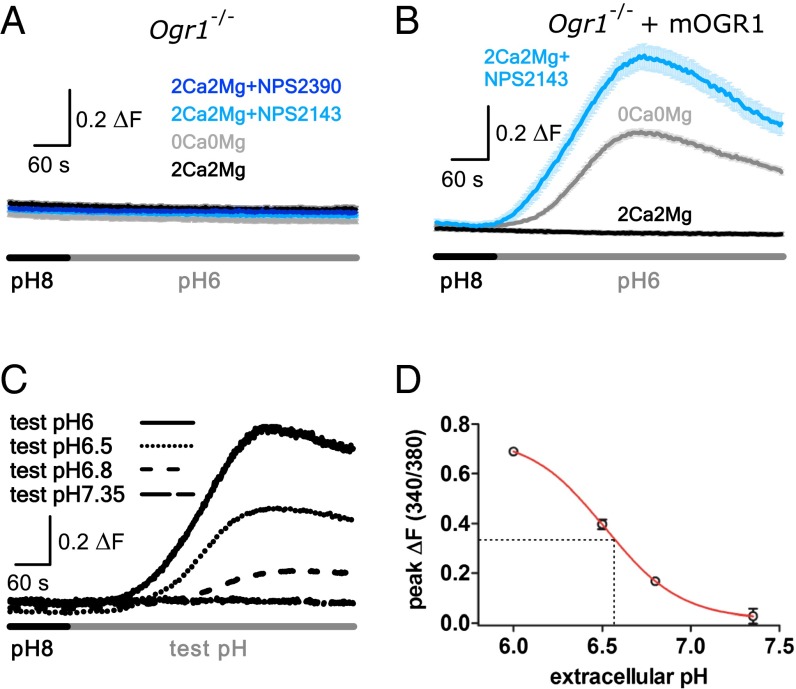
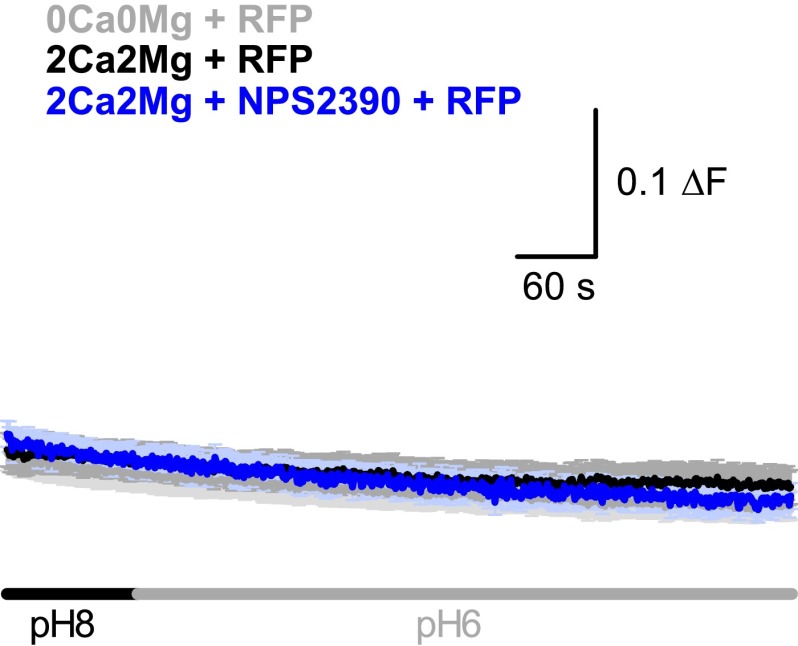
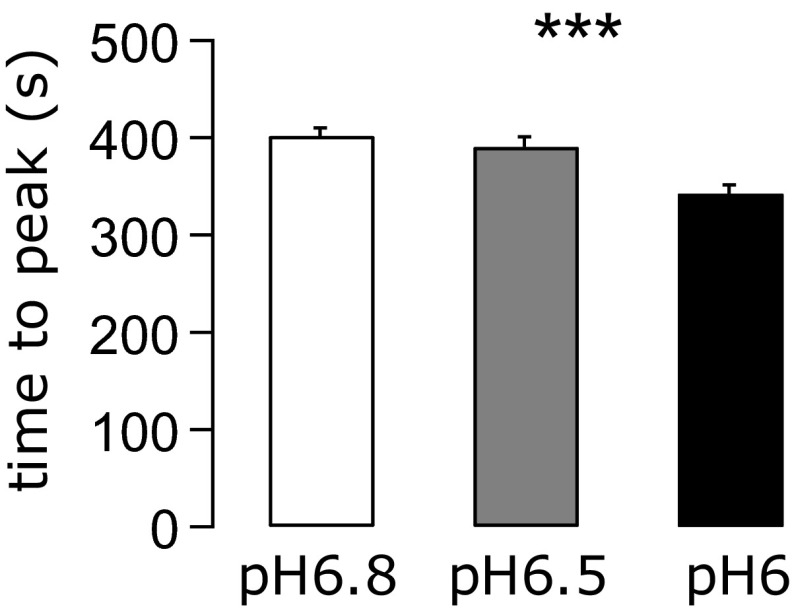
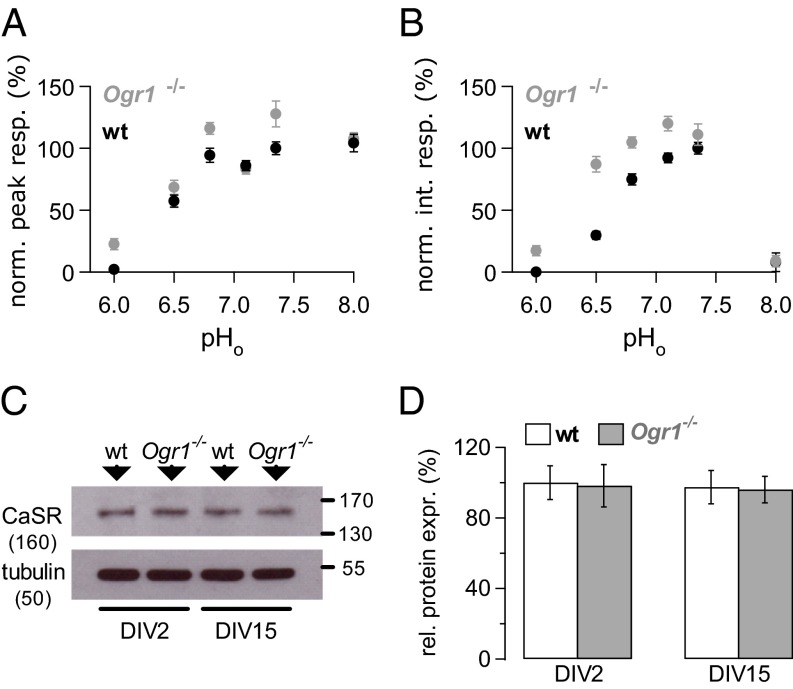
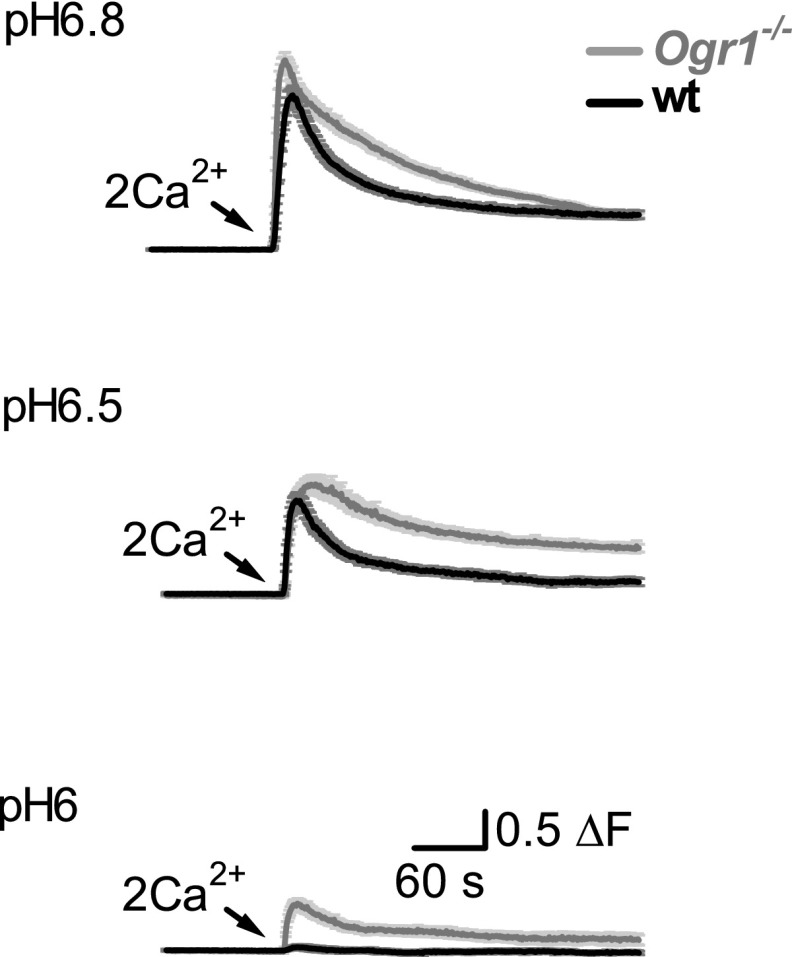
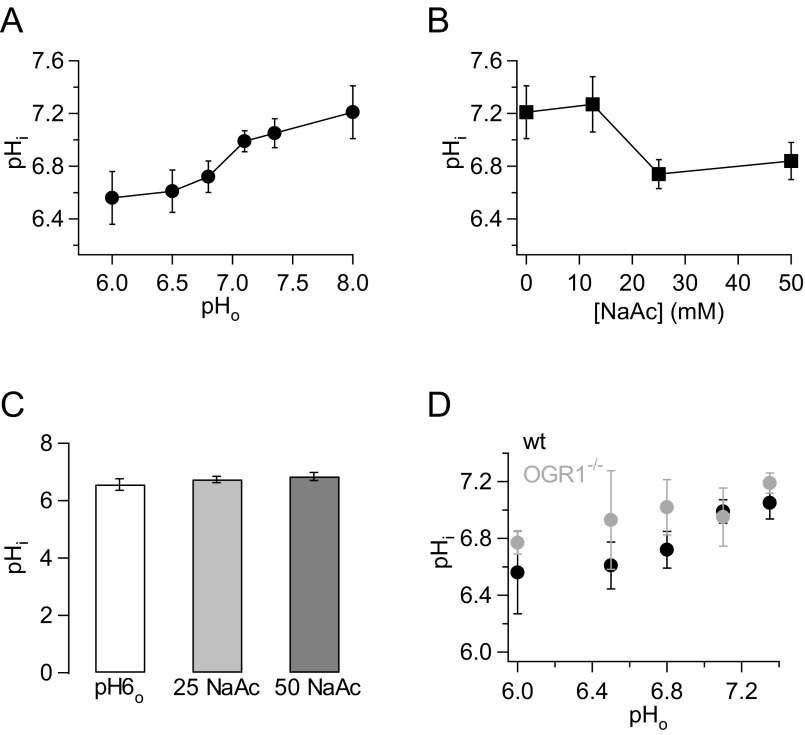
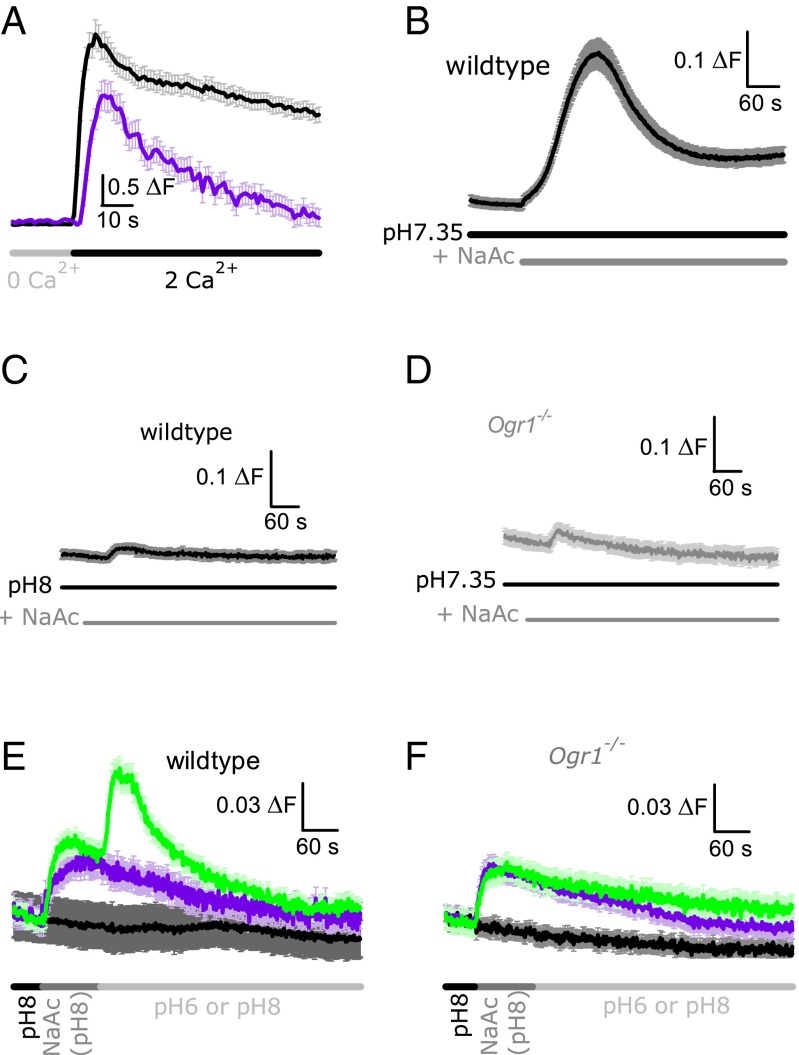
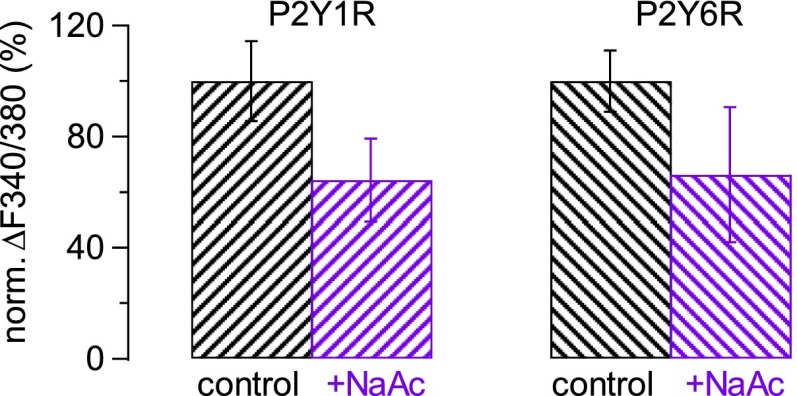
References
-
- Hussell T, Bell TJ. Alveolar macrophages: Plasticity in a tissue-specific context. Nat Rev Immunol. 2014;14(2):81–93. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
