

A DEMETER-like DNA demethylase governs tomato fruit ripening
- PMID: 26261318
- PMCID: PMC4553810
- DOI: 10.1073/pnas.1503362112
A DEMETER-like DNA demethylase governs tomato fruit ripening
Abstract
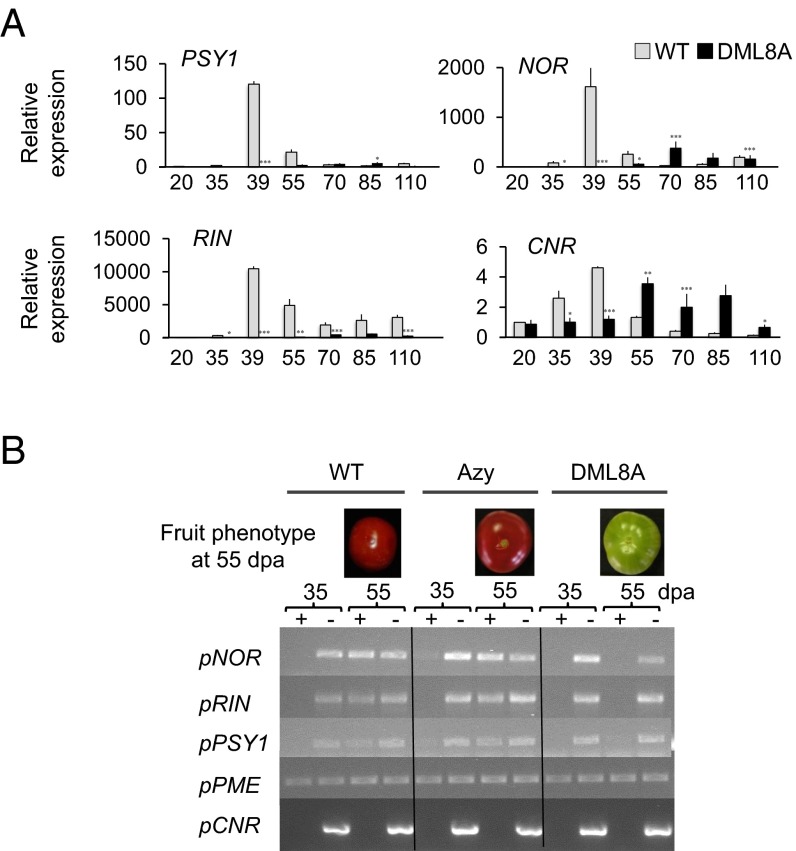
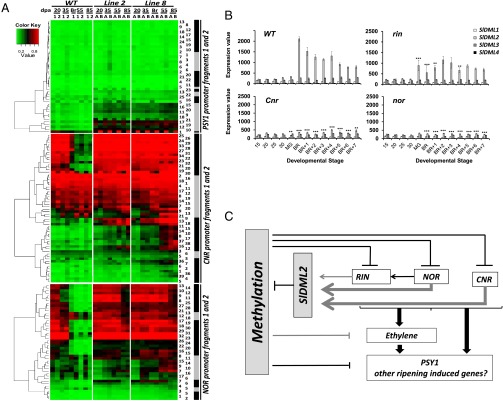
In plants, genomic DNA methylation which contributes to development and stress responses can be actively removed by DEMETER-like DNA demethylases (DMLs). Indeed, in Arabidopsis DMLs are important for maternal imprinting and endosperm demethylation, but only a few studies demonstrate the developmental roles of active DNA demethylation conclusively in this plant. Here, we show a direct cause and effect relationship between active DNA demethylation mainly mediated by the tomato DML, SlDML2, and fruit ripening- an important developmental process unique to plants. RNAi SlDML2 knockdown results in ripening inhibition via hypermethylation and repression of the expression of genes encoding ripening transcription factors and rate-limiting enzymes of key biochemical processes such as carotenoid synthesis. Our data demonstrate that active DNA demethylation is central to the control of ripening in tomato.
Keywords: DNA glycosylase lyase; active DNA demethylation; epigenetic; fruit ripening; tomato.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Saze H, Tsugane K, Kanno T, Nishimura T. DNA methylation in plants: Relationship to small RNAs and histone modifications, and functions in transposon inactivation. Plant Cell Physiol. 2012;53(5):766–784. - PubMed
-
- Chan SW, Henderson IR, Jacobsen SE. Gardening the genome: DNA methylation in Arabidopsis thaliana. Nat Rev Genet. 2005;6(5):351–360. - PubMed
-
- Zhang M, Kimatu JN, Xu K, Liu B. DNA cytosine methylation in plant development. J Genet Genomics. 2010;37(1):1–12. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
