

Early evolution of the angiosperm clade Asteraceae in the Cretaceous of Antarctica
- PMID: 26261324
- PMCID: PMC4568267
- DOI: 10.1073/pnas.1423653112
Early evolution of the angiosperm clade Asteraceae in the Cretaceous of Antarctica
Abstract
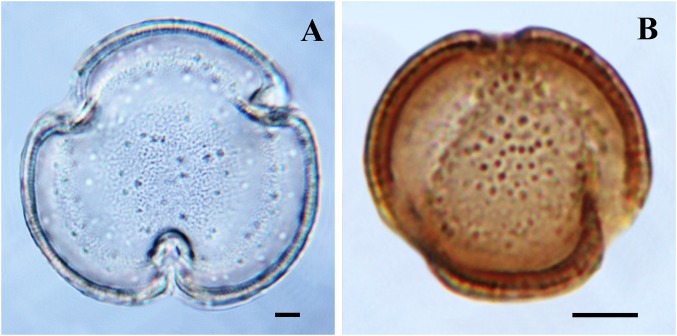
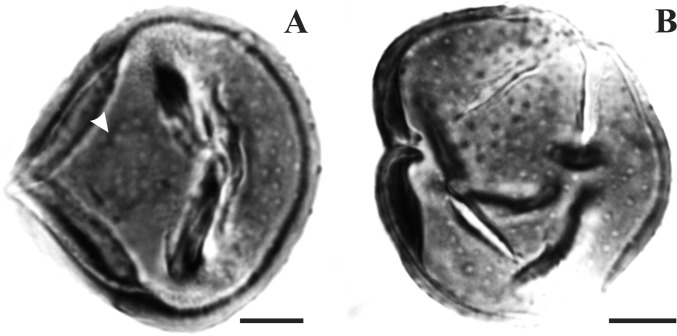
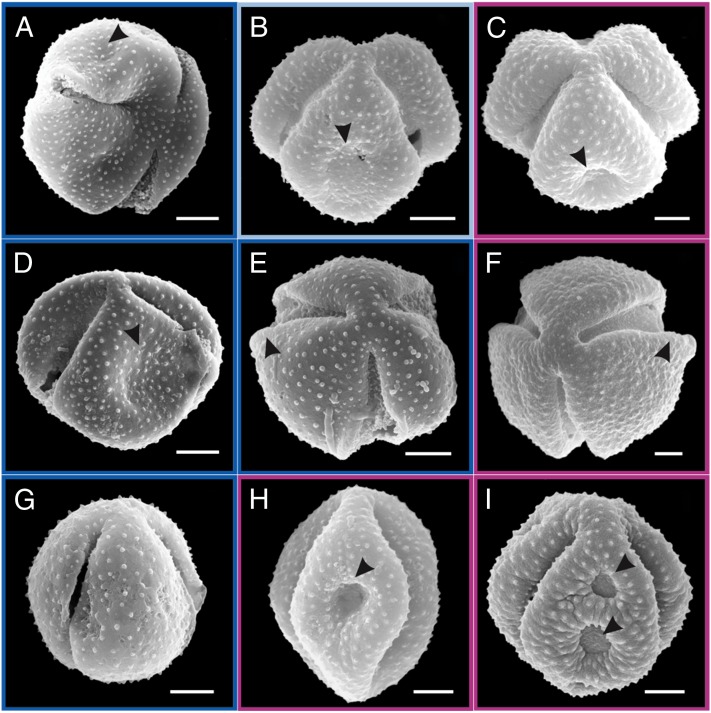
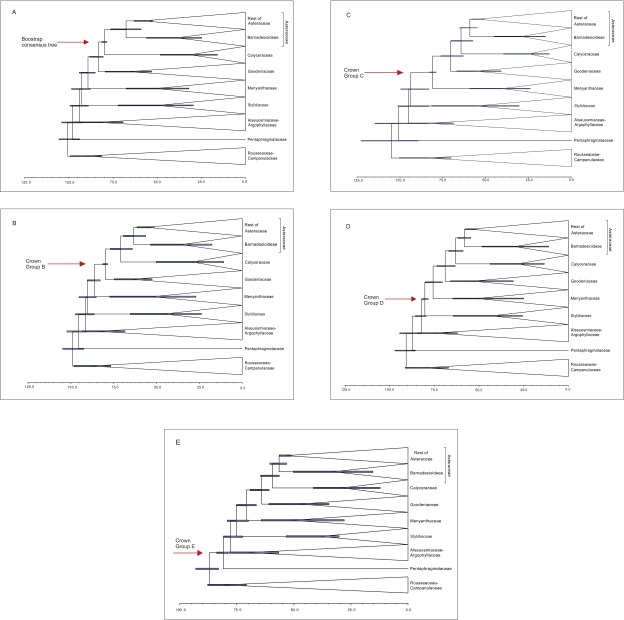
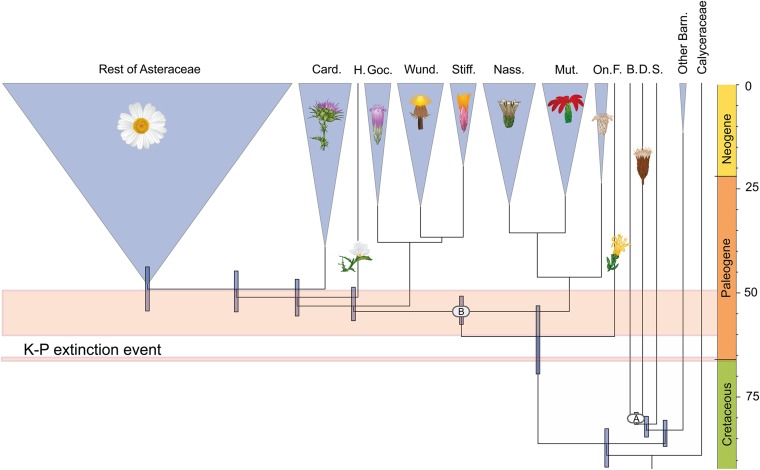
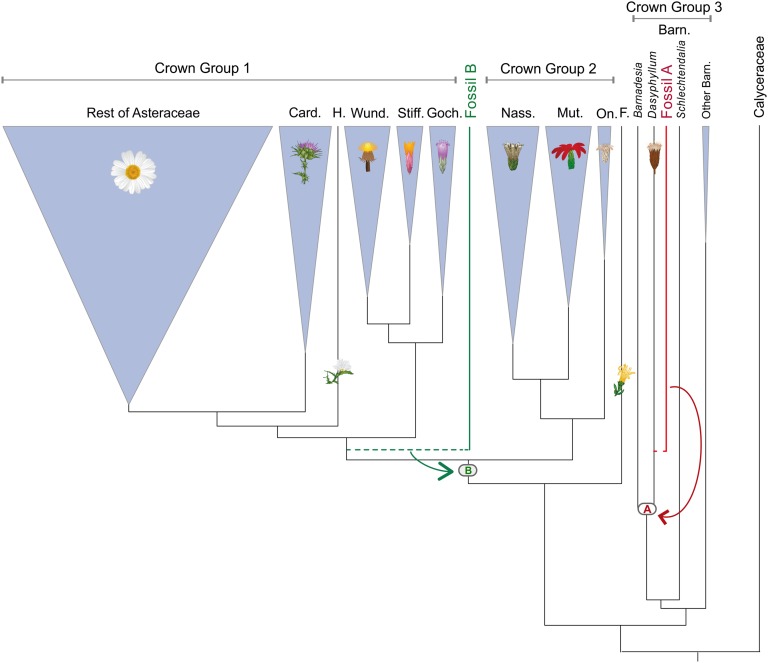
The Asteraceae (sunflowers and daisies) are the most diverse family of flowering plants. Despite their prominent role in extant terrestrial ecosystems, the early evolutionary history of this family remains poorly understood. Here we report the discovery of a number of fossil pollen grains preserved in dinosaur-bearing deposits from the Late Cretaceous of Antarctica that drastically pushes back the timing of assumed origin of the family. Reliably dated to ∼76-66 Mya, these specimens are about 20 million years older than previously known records for the Asteraceae. Using a phylogenetic approach, we interpreted these fossil specimens as members of an extinct early diverging clade of the family, associated with subfamily Barnadesioideae. Based on a molecular phylogenetic tree calibrated using fossils, including the ones reported here, we estimated that the most recent common ancestor of the family lived at least 80 Mya in Gondwana, well before the thermal and biogeographical isolation of Antarctica. Most of the early diverging lineages of the family originated in a narrow time interval after the K/P boundary, 60-50 Mya, coinciding with a pronounced climatic warming during the Late Paleocene and Early Eocene, and the scene of a dramatic rise in flowering plant diversity. Our age estimates reduce earlier discrepancies between the age of the fossil record and previous molecular estimates for the origin of the family, bearing important implications in the evolution of flowering plants in general.
Keywords: Antarctica; Asteraceae; evolution; fossil; phylogenetics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Comment in
-
Reply to Panero: Robust phylogenetic placement of fossil pollen grains: The case of Asteraceae.Proc Natl Acad Sci U S A. 2016 Jan 26;113(4):E412. doi: 10.1073/pnas.1521642113. Epub 2015 Dec 23. Proc Natl Acad Sci U S A. 2016. PMID: 26699489 Free PMC article. No abstract available.
-
Phylogenetic uncertainty and fossil calibration of Asteraceae chronograms.Proc Natl Acad Sci U S A. 2016 Jan 26;113(4):E411. doi: 10.1073/pnas.1517649113. Epub 2015 Dec 23. Proc Natl Acad Sci U S A. 2016. PMID: 26699490 Free PMC article. No abstract available.
References
-
- Friis EM, Crane PR, Pedersen KR. Early Flowers and Angiosperm Evolution. Cambridge Univ Press; Cambridge, UK: 2011.
-
- Funk VA, et al. Compositae metatrees: The next generation. In: Funk VA, Susanna A, Stuessy TF, Bayer RJ, editors. Systematics, Evolution, and Biogeography of Compositae. IAPT; Vienna, Austria: 2009. pp. 747–777.
-
- Barreda VD, et al. Eocene Patagonia fossils of the daisy family. Science. 2010;329(5999):1621. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
