

Multilayered Regulation of Ethylene Induction Plays a Positive Role in Arabidopsis Resistance against Pseudomonas syringae
- PMID: 26265775
- PMCID: PMC4577408
- DOI: 10.1104/pp.15.00659
Multilayered Regulation of Ethylene Induction Plays a Positive Role in Arabidopsis Resistance against Pseudomonas syringae
Abstract
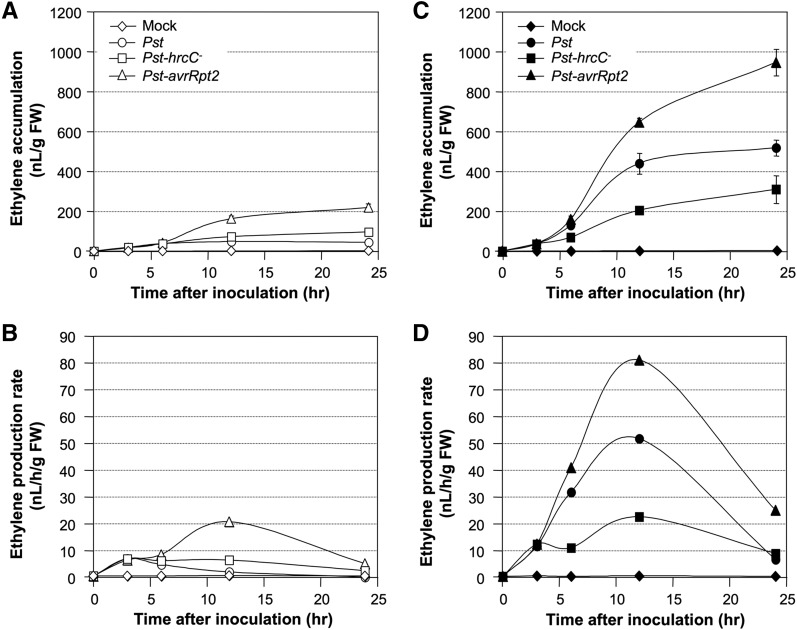
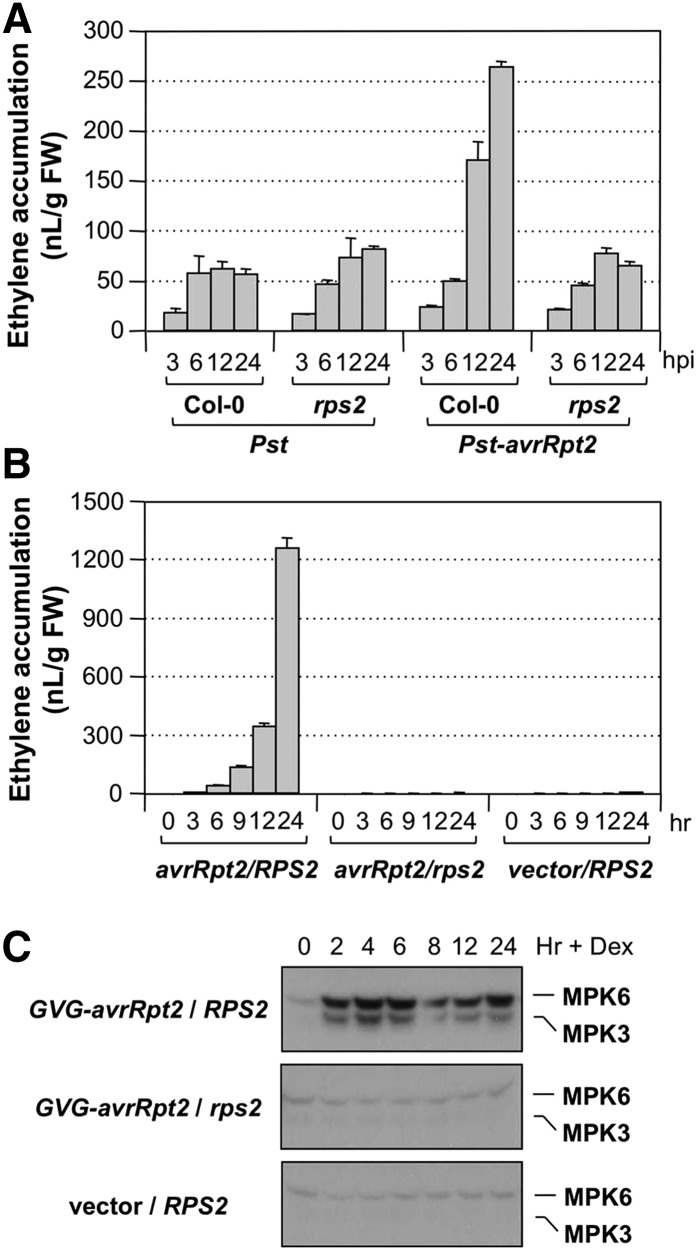
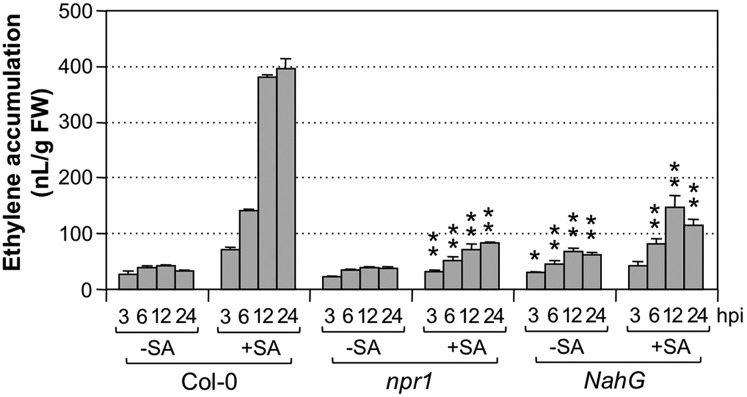
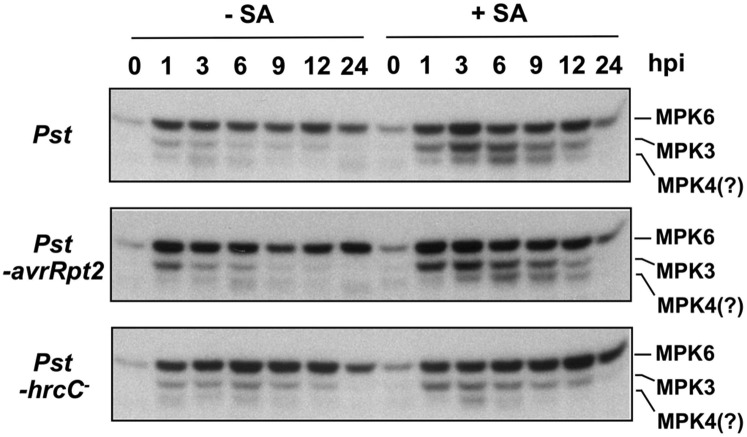
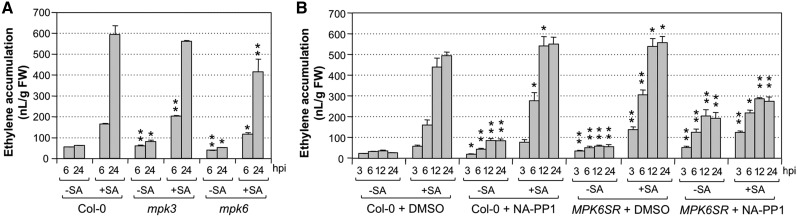
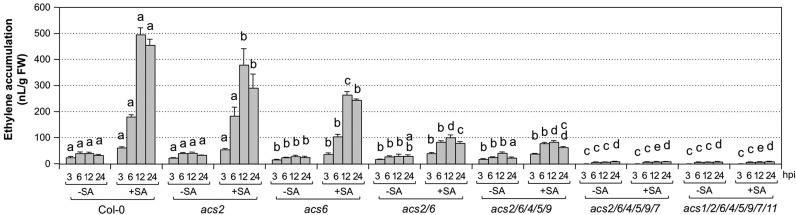
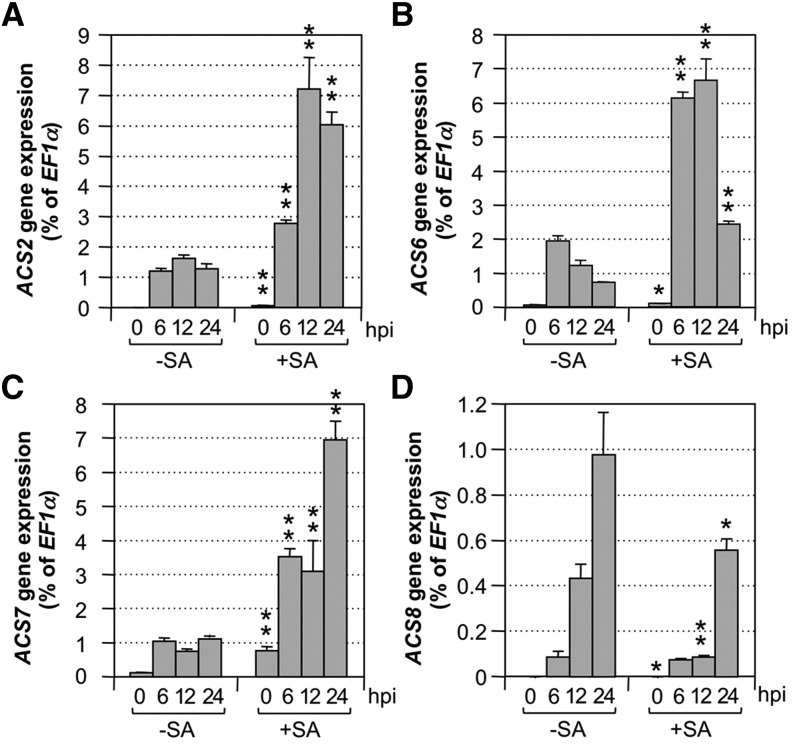
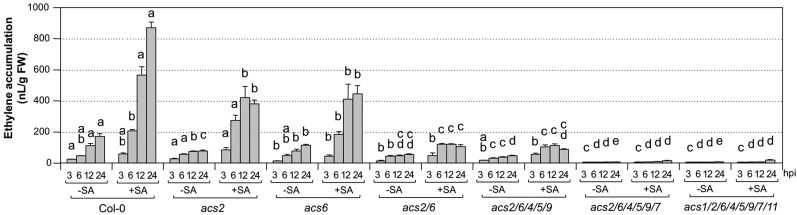
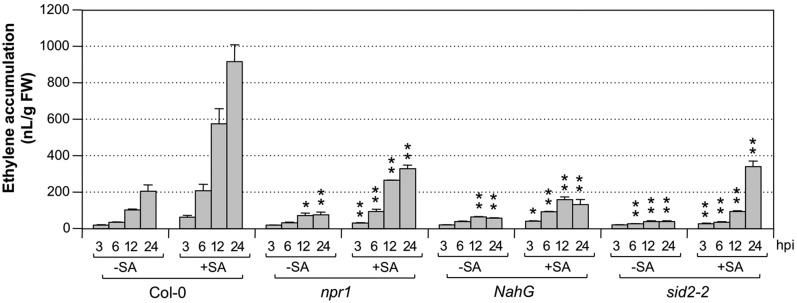
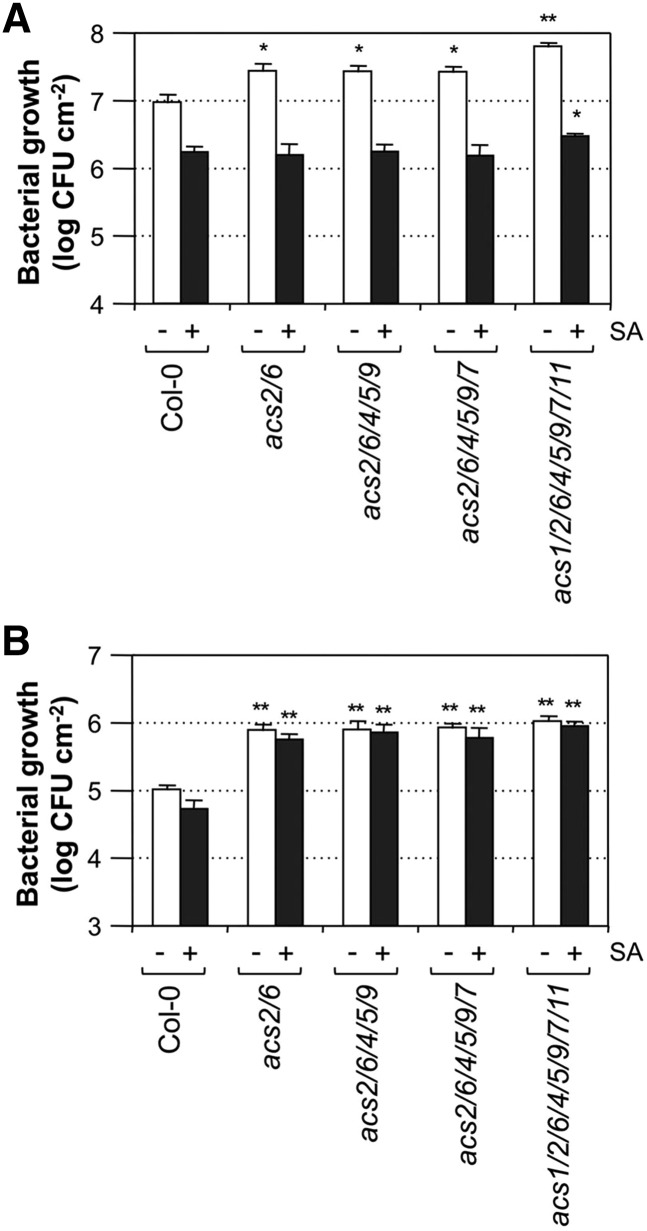
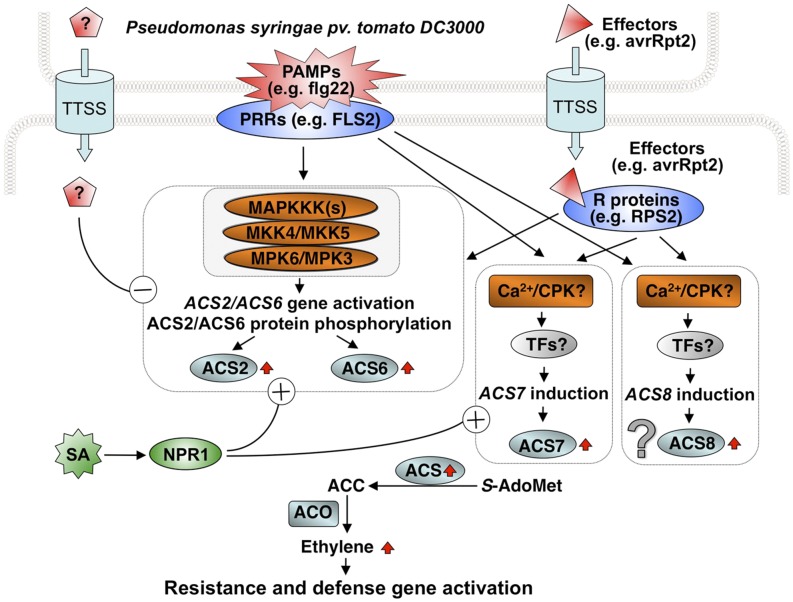
Ethylene, a key phytohormone involved in plant-pathogen interaction, plays a positive role in plant resistance against fungal pathogens. However, its function in plant bacterial resistance remains unclear. Here, we report a detailed analysis of ethylene induction in Arabidopsis (Arabidopsis thaliana) in response to Pseudomonas syringae pv tomato DC3000 (Pst). Ethylene biosynthesis is highly induced in both pathogen/microbe-associated molecular pattern (PAMP)-triggered immunity and effector-triggered immunity (ETI), and the induction is potentiated by salicylic acid (SA) pretreatment. In addition, Pst actively suppresses PAMP-triggered ethylene induction in a type III secretion system-dependent manner. SA potentiation of ethylene induction is dependent mostly on MITOGEN-ACTIVATED PROTEIN KINASE6 (MPK6) and MPK3 and their downstream ACS2 and ACS6, two type I isoforms of 1-aminocyclopropane-1-carboxylic acid synthases (ACSs). ACS7, a type III ACS whose expression is enhanced by SA pretreatment, is also involved. Pst expressing the avrRpt2 effector gene (Pst-avrRpt2), which is capable of triggering ETI, induces a higher level of ethylene production, and the elevated portion is dependent on SALICYLIC ACID INDUCTION DEFICIENT2 and NONEXPRESSER OF PATHOGENESIS-RELATED GENE1, two key players in SA biosynthesis and signaling. High-order ACS mutants with reduced ethylene induction are more susceptible to both Pst and Pst-avrRpt2, demonstrating a positive role of ethylene in plant bacterial resistance mediated by both PAMP-triggered immunity and ETI.
© 2015 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Alfano JR, Collmer A (2004) Type III secretion system effector proteins: double agents in bacterial disease and plant defense. Annu Rev Phytopathol 42: 385–414 - PubMed
-
- An C, Mou Z (2011) Salicylic acid and its function in plant immunity. J Integr Plant Biol 53: 412–428 - PubMed
-
- Anderson JP, Badruzsaufari E, Schenk PM, Manners JM, Desmond OJ, Ehlert C, Maclean DJ, Ebert PR, Kazan K (2004) Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell 16: 3460–3479 - PMC - PubMed
-
- Ausubel FM. (2005) Are innate immune signaling pathways in plants and animals conserved? Nat Immunol 6: 973–979 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
