

Spatiotemporal Spike Coding of Behavioral Adaptation in the Dorsal Anterior Cingulate Cortex
- PMID: 26266537
- PMCID: PMC4534466
- DOI: 10.1371/journal.pbio.1002222
Spatiotemporal Spike Coding of Behavioral Adaptation in the Dorsal Anterior Cingulate Cortex
Abstract
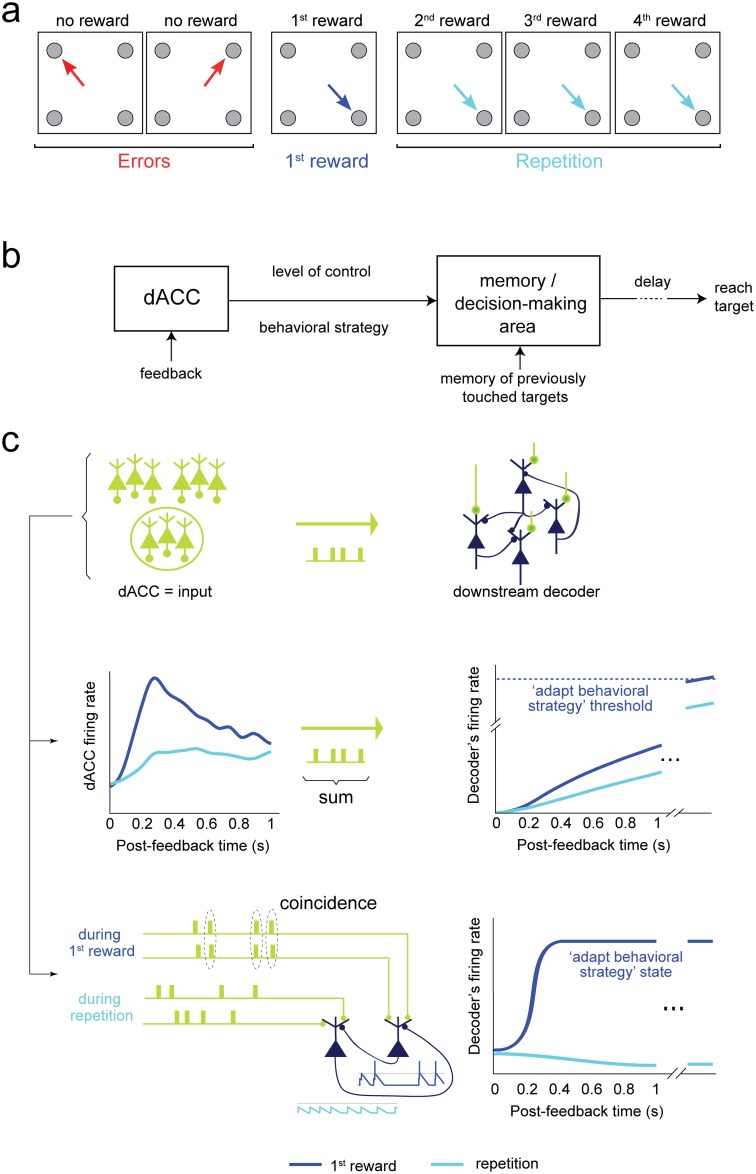
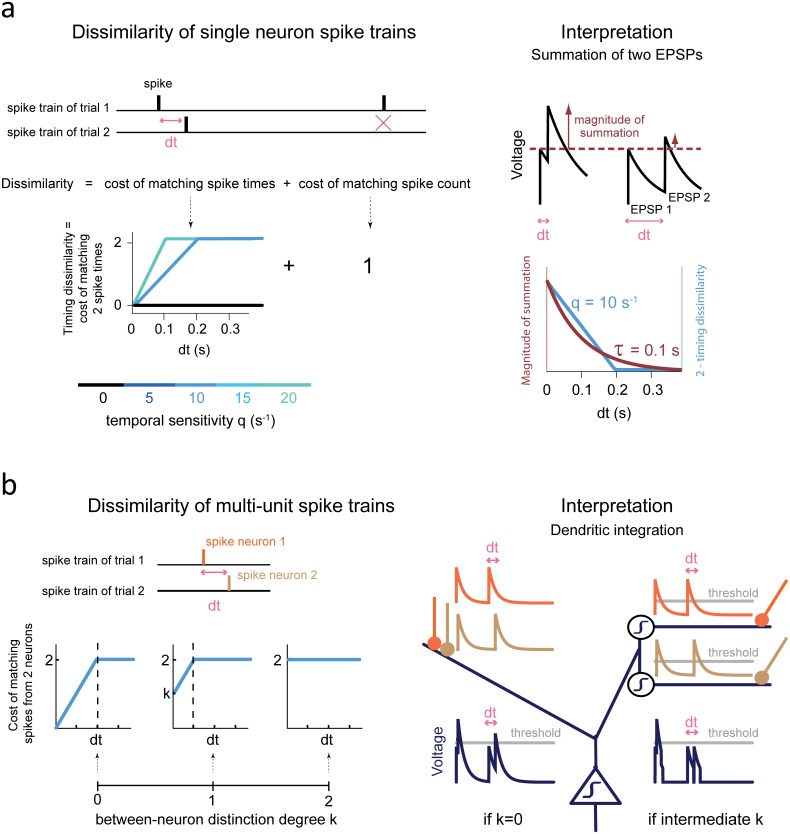
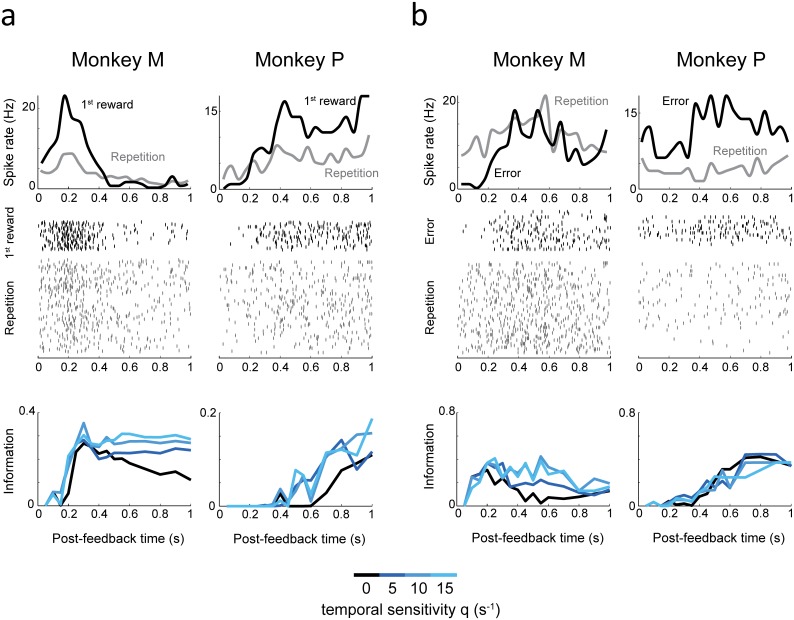
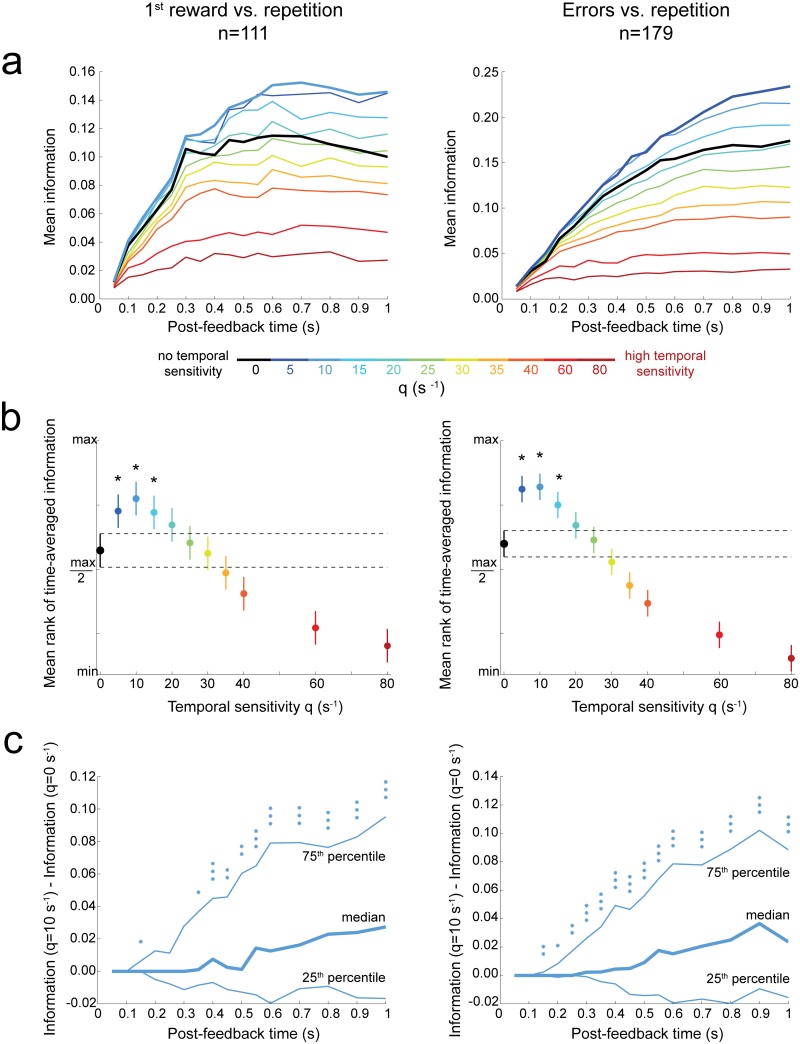
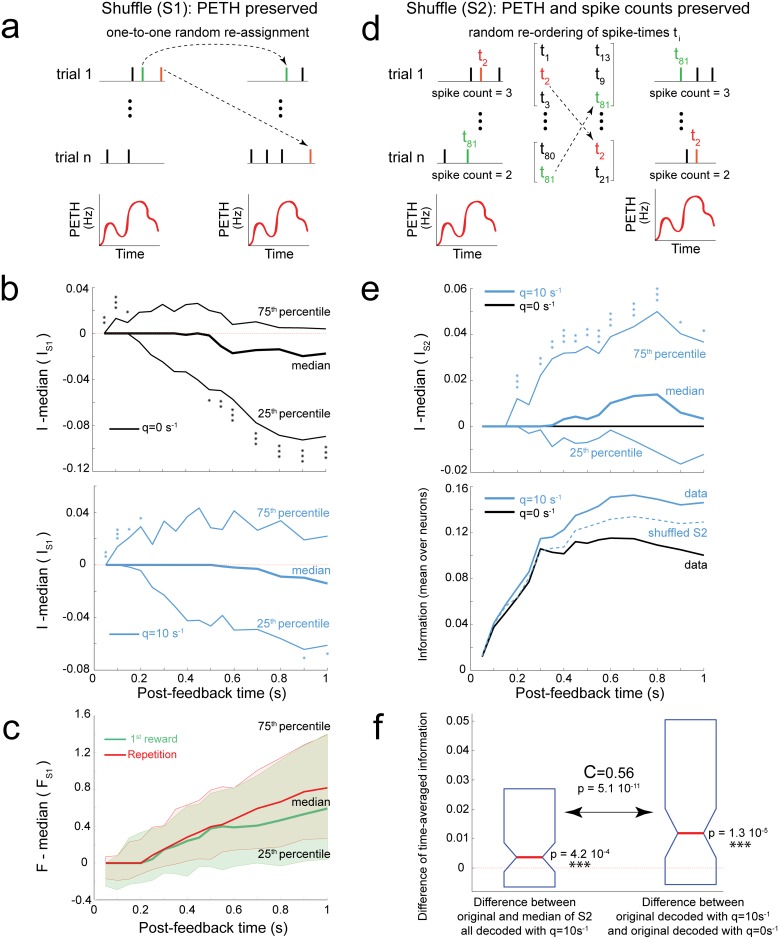
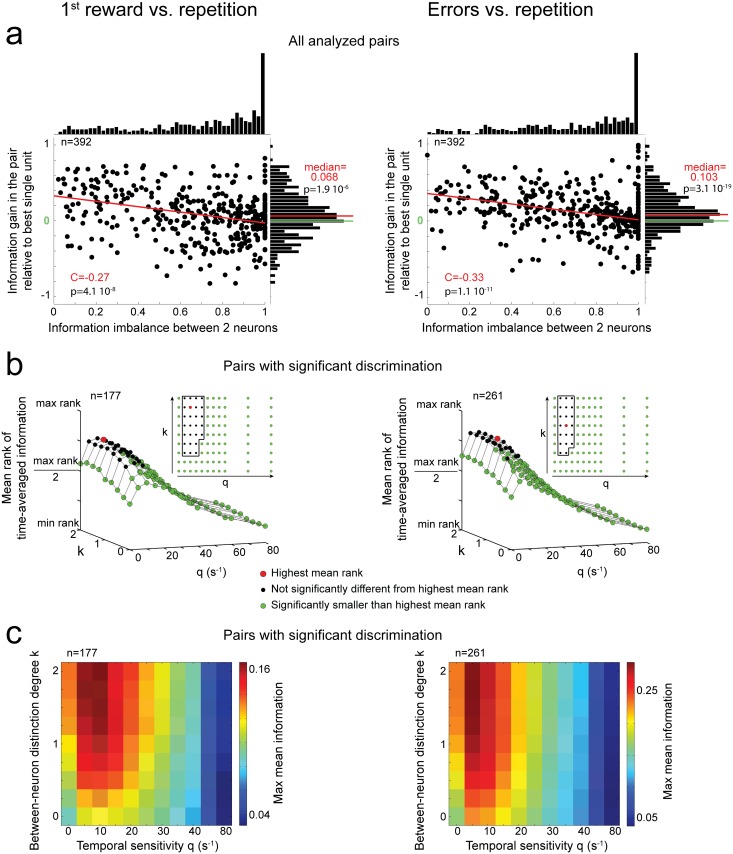
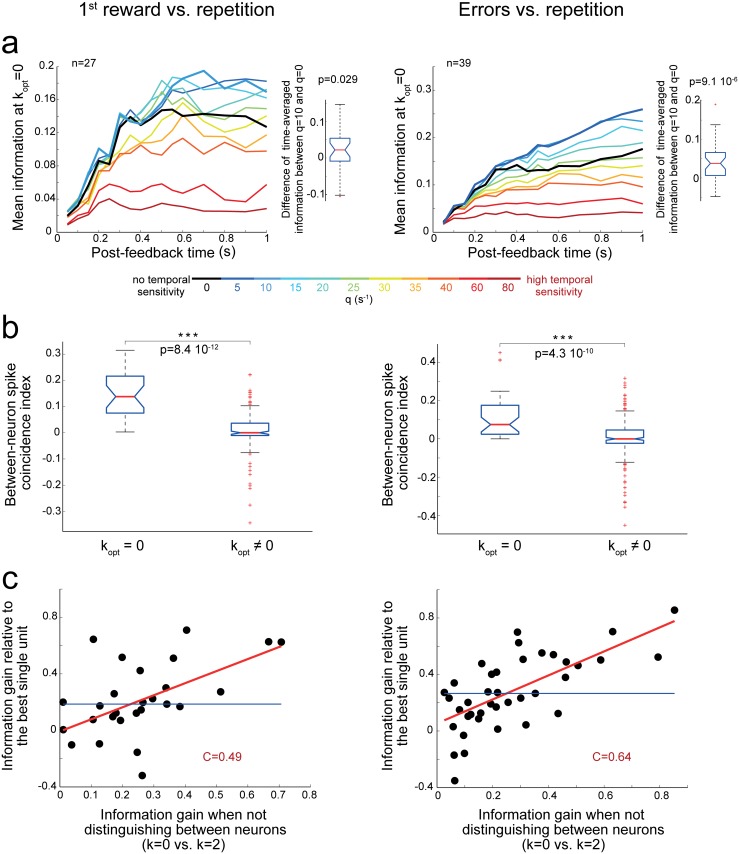
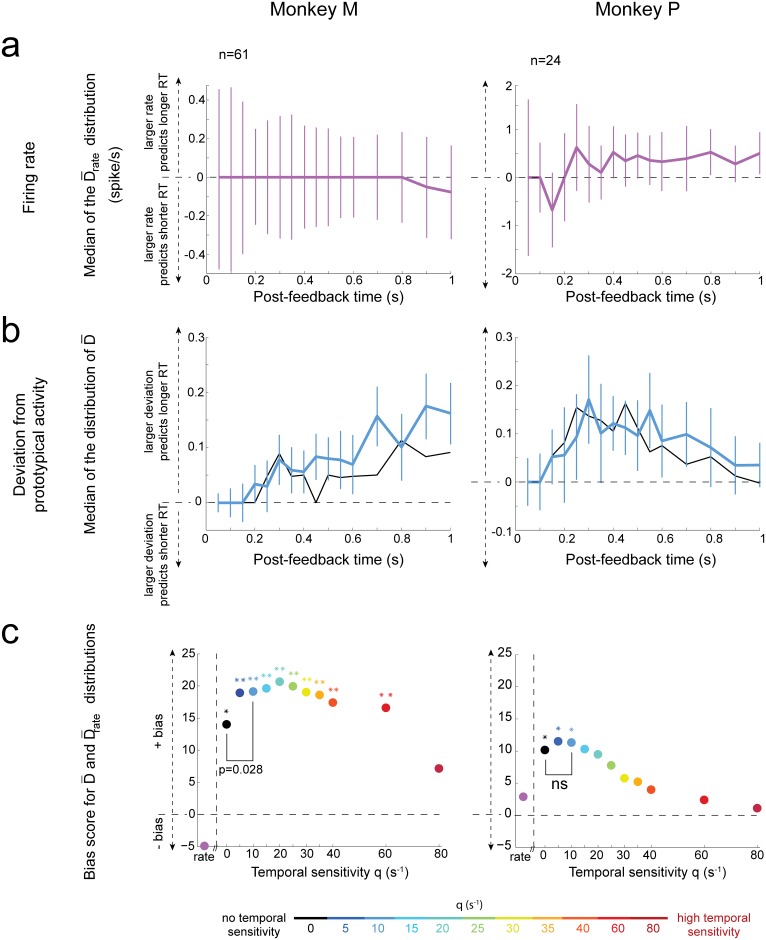
The frontal cortex controls behavioral adaptation in environments governed by complex rules. Many studies have established the relevance of firing rate modulation after informative events signaling whether and how to update the behavioral policy. However, whether the spatiotemporal features of these neuronal activities contribute to encoding imminent behavioral updates remains unclear. We investigated this issue in the dorsal anterior cingulate cortex (dACC) of monkeys while they adapted their behavior based on their memory of feedback from past choices. We analyzed spike trains of both single units and pairs of simultaneously recorded neurons using an algorithm that emulates different biologically plausible decoding circuits. This method permits the assessment of the performance of both spike-count and spike-timing sensitive decoders. In response to the feedback, single neurons emitted stereotypical spike trains whose temporal structure identified informative events with higher accuracy than mere spike count. The optimal decoding time scale was in the range of 70-200 ms, which is significantly shorter than the memory time scale required by the behavioral task. Importantly, the temporal spiking patterns of single units were predictive of the monkeys' behavioral response time. Furthermore, some features of these spiking patterns often varied between jointly recorded neurons. All together, our results suggest that dACC drives behavioral adaptation through complex spatiotemporal spike coding. They also indicate that downstream networks, which decode dACC feedback signals, are unlikely to act as mere neural integrators.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Brunel N, Wang XJ. Effects of neuromodulation in a cortical network model of object working memory dominated by recurrent inhibition. Journal of computational neuroscience. 2001. Jul-Aug;11(1):63–85. - PubMed
-
- Mongillo G, Barak O, Tsodyks M. Synaptic theory of working memory. Science (New York, NY. 2008. March 14;319(5869):1543–6. - PubMed
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
