

Mutations Proximal to Sites of Autoproteolysis and the α-Helix That Co-evolve under Drug Pressure Modulate the Autoprocessing and Vitality of HIV-1 Protease
- PMID: 26266692
- PMCID: PMC5590646
- DOI: 10.1021/acs.biochem.5b00759
Mutations Proximal to Sites of Autoproteolysis and the α-Helix That Co-evolve under Drug Pressure Modulate the Autoprocessing and Vitality of HIV-1 Protease
Abstract
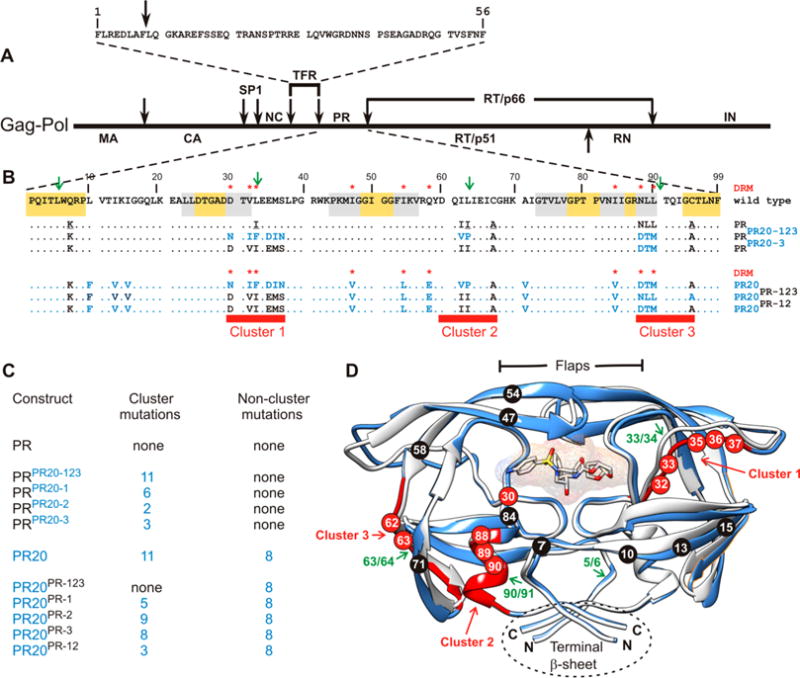
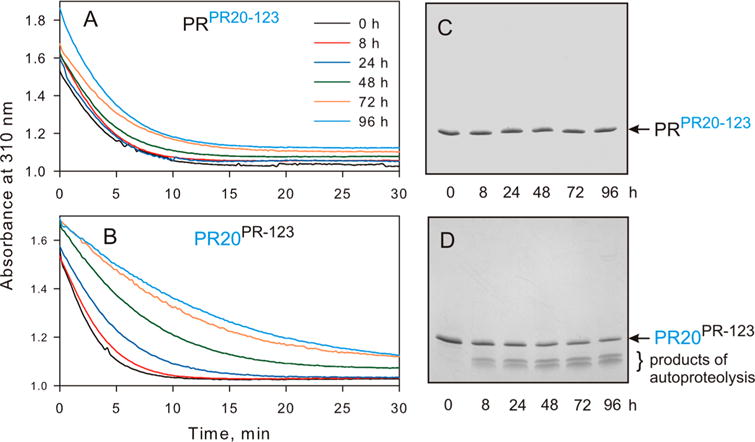
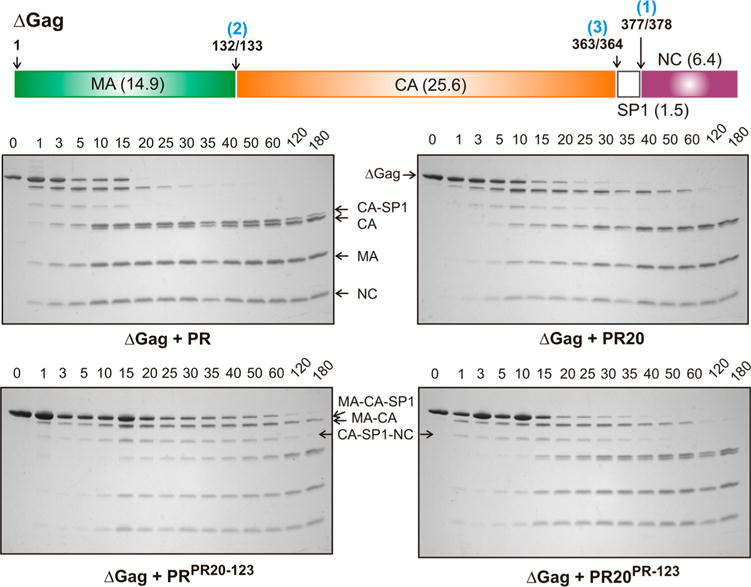
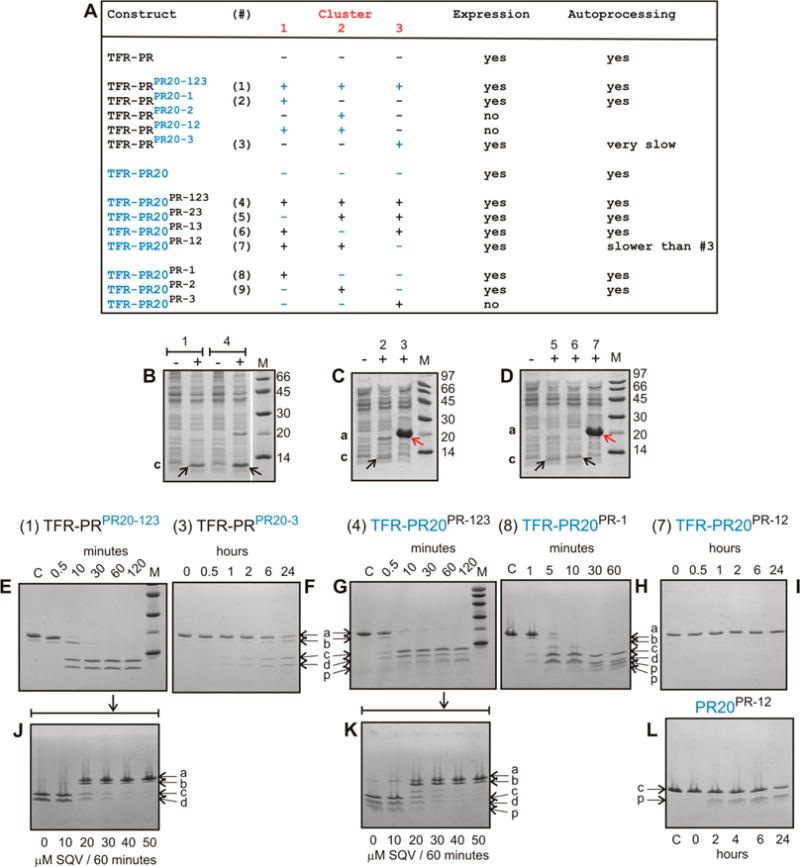
N-Terminal self-cleavage (autoprocessing) of the HIV-1 protease precursor is crucial for liberating the active dimer. Under drug pressure, evolving mutations are predicted to modulate autoprocessing, and the reduced catalytic activity of the mature protease (PR) is likely compensated by enhanced conformational/dimer stability and reduced susceptibility to self-degradation (autoproteolysis). One such highly evolved, multidrug resistant protease, PR20, bears 19 mutations contiguous to sites of autoproteolysis in retroviral proteases, namely clusters 1-3 comprising residues 30-37, 60-67, and 88-95, respectively, accounting for 11 of the 19 mutations. By systematically replacing corresponding clusters in PR with those of PR20, and vice versa, we assess their influence on the properties mentioned above and observe no strict correlation. A 10-35-fold decrease in the cleavage efficiency of peptide substrates by PR20, relative to PR, is reflected by an only ∼4-fold decrease in the rate of Gag processing with no change in cleavage order. Importantly, optimal N-terminal autoprocessing requires all 19 PR20 mutations as evaluated in vitro using the model precursor TFR-PR20 in which PR is flanked by the transframe region. Substituting PR20 cluster 3 into TFR-PR (TFR-PR(PR20-3)) requires the presence of PR20 cluster 1 and/or 2 for autoprocessing. In accordance, substituting PR clusters 1 and 2 into TFR-PR20 affects the rate of autoprocessing more drastically (>300-fold) compared to that of TFR-PR(PR20-3) because of the cumulative effect of eight noncluster mutations present in TFR-PR20(PR-12). Overall, these studies imply that drug resistance involves a complex synchronized selection of mutations modulating all of the properties mentioned above governing PR regulation and function.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Louis JM, Weber IT, Tozser J, Marius Clore G, Gronenborn AM. HIV-1 protease: maturation, enzyme specificity, and drug resistance. Adv Pharmacol. 2000;49:111–146. - PubMed
-
- Louis JM, Ishima R, Torchia DA, Weber IT. HIV-1 protease: structure, dynamics, and inhibition. Adv Pharmacol. 2007;55:261–298. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
