

A phosphoinositide-binding cluster in cavin1 acts as a molecular sensor for cavin1 degradation
- PMID: 26269585
- PMCID: PMC4603927
- DOI: 10.1091/mbc.E15-06-0359
A phosphoinositide-binding cluster in cavin1 acts as a molecular sensor for cavin1 degradation
Abstract
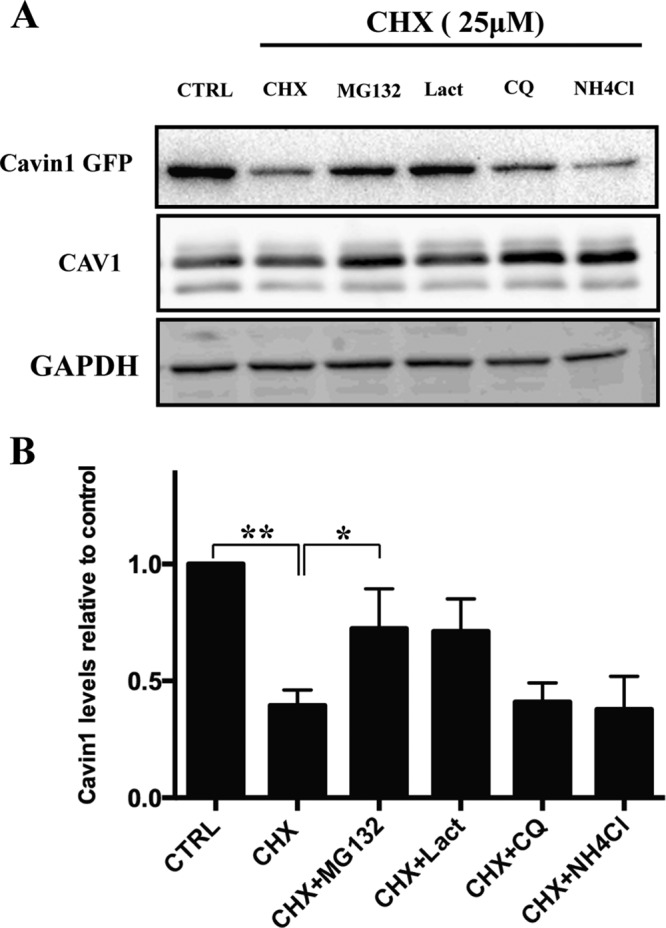
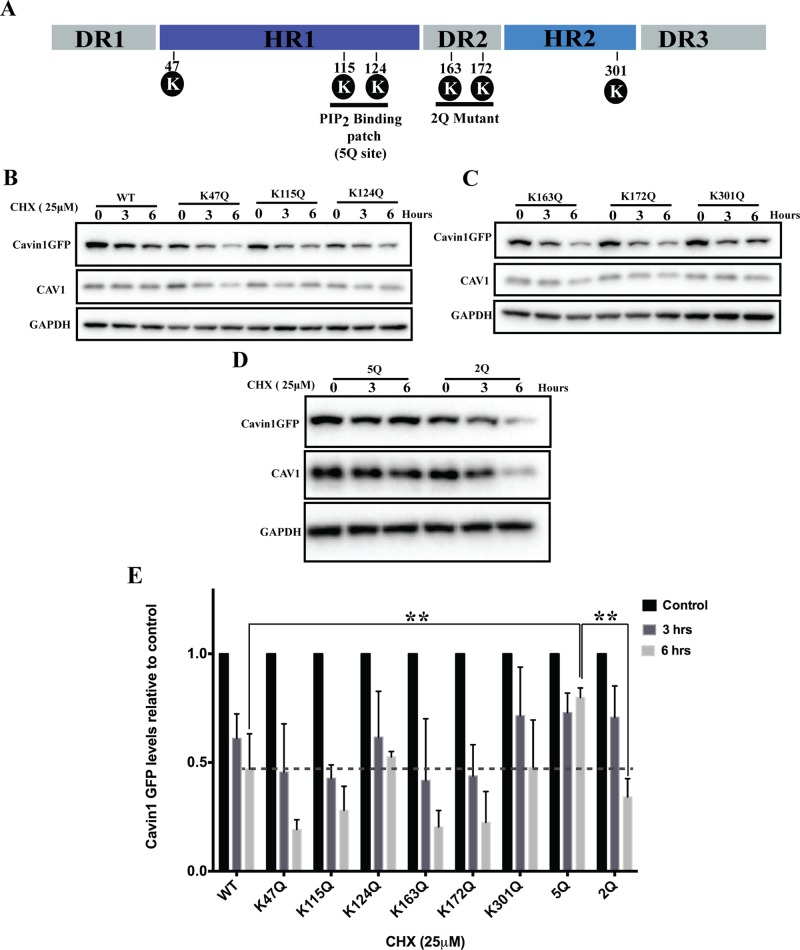
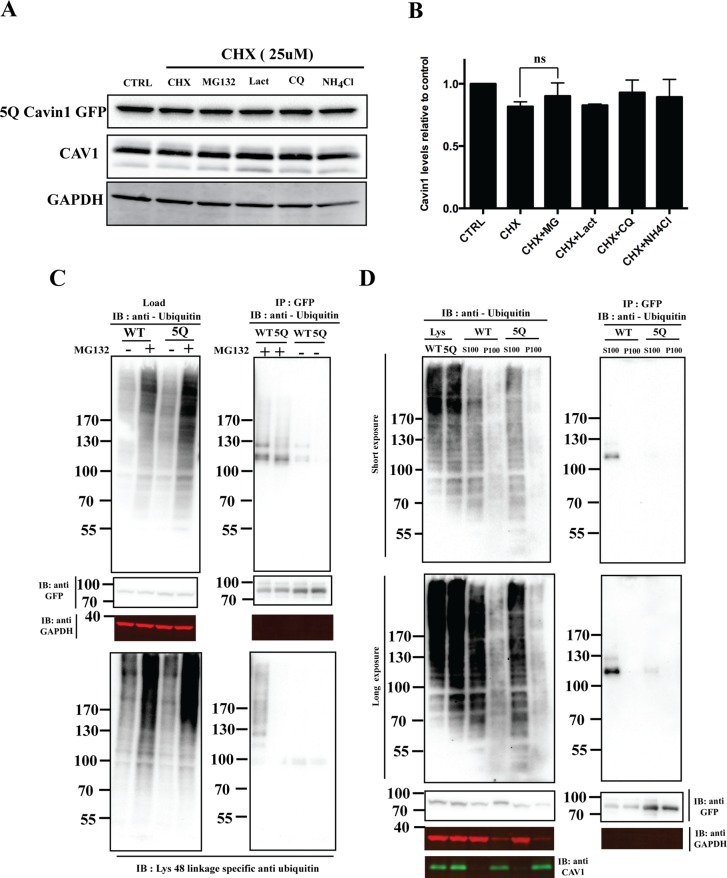
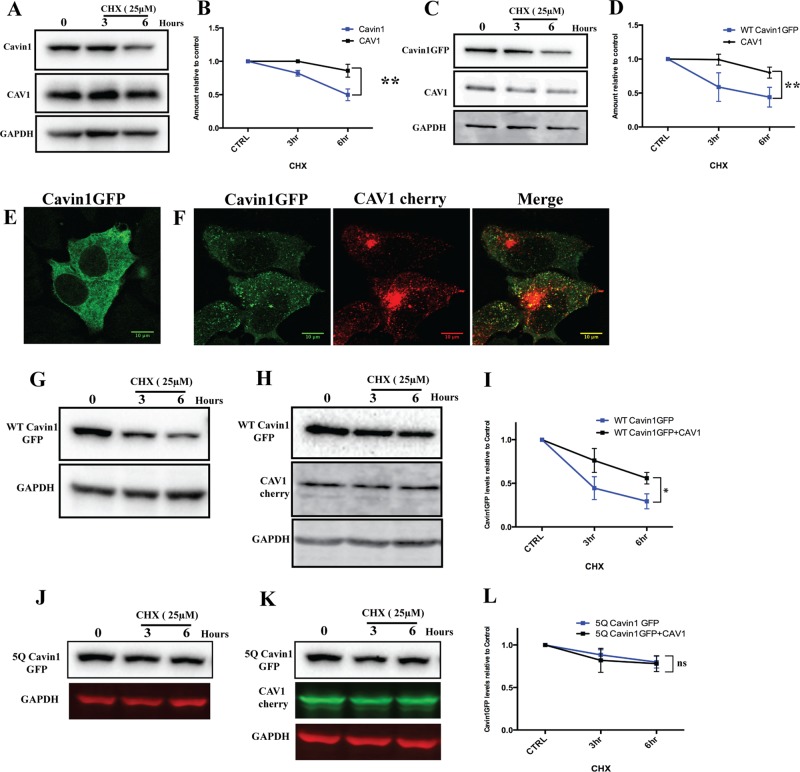
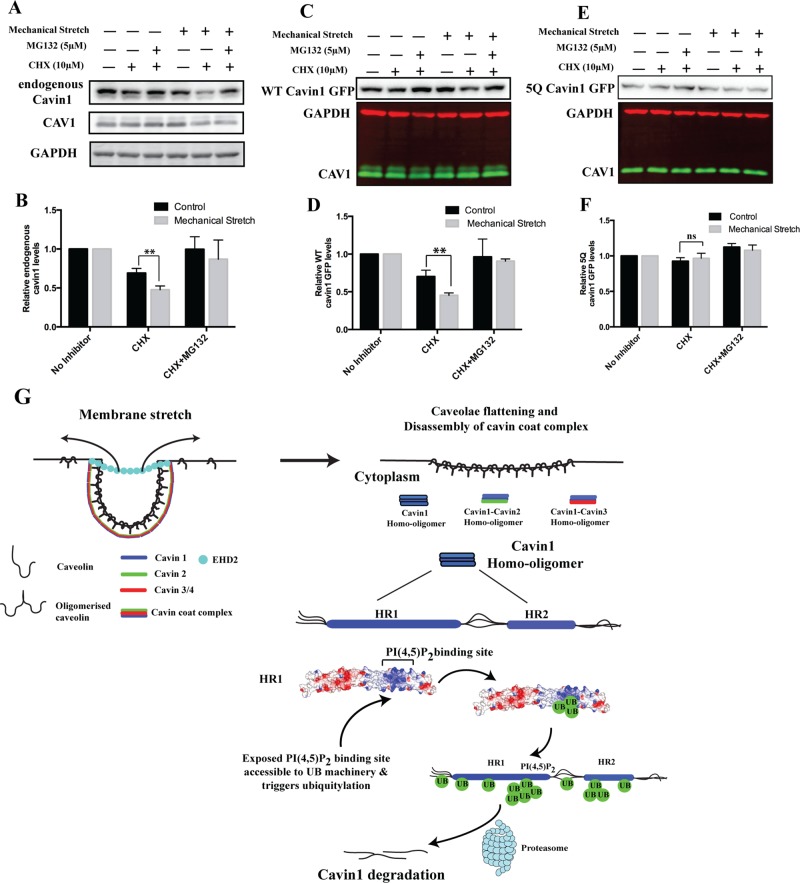
Caveolae are abundant surface organelles implicated in a range of cellular processes. Two classes of proteins work together to generate caveolae: integral membrane proteins termed caveolins and cytoplasmic coat proteins called cavins. Caveolae respond to membrane stress by releasing cavins into the cytosol. A crucial aspect of this model is tight regulation of cytosolic pools of cavin under resting conditions. We now show that a recently identified region of cavin1 that can bind phosphoinositide (PI) lipids is also a major site of ubiquitylation. Ubiquitylation of lysines within this site leads to rapid proteasomal degradation. In cells that lack caveolins and caveolae, cavin1 is cytosolic and rapidly degraded as compared with cells in which cavin1 is associated with caveolae. Membrane stretching causes caveolar disassembly, release of cavin complexes into the cytosol, and increased proteasomal degradation of wild-type cavin1 but not mutant cavin1 lacking the major ubiquitylation site. Release of cavin1 from caveolae thus leads to exposure of key lysine residues in the PI-binding region, acting as a trigger for cavin1 ubiquitylation and down-regulation. This mutually exclusive PI-binding/ubiquitylation mechanism may help maintain low levels of cytosolic cavin1 in resting cells, a prerequisite for cavins acting as signaling modules following release from caveolae.
© 2015 Tillu et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
