

A Comparison of Ci/Gli Activity as Regulated by Sufu in Drosophila and Mammalian Hedgehog Response
- PMID: 26271100
- PMCID: PMC4536226
- DOI: 10.1371/journal.pone.0135804
A Comparison of Ci/Gli Activity as Regulated by Sufu in Drosophila and Mammalian Hedgehog Response
Abstract
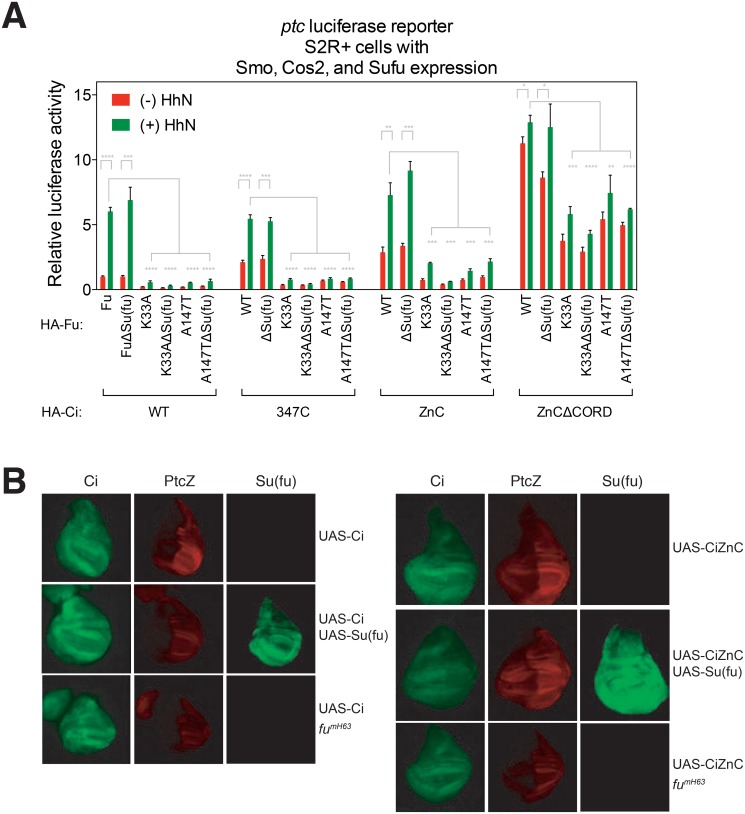
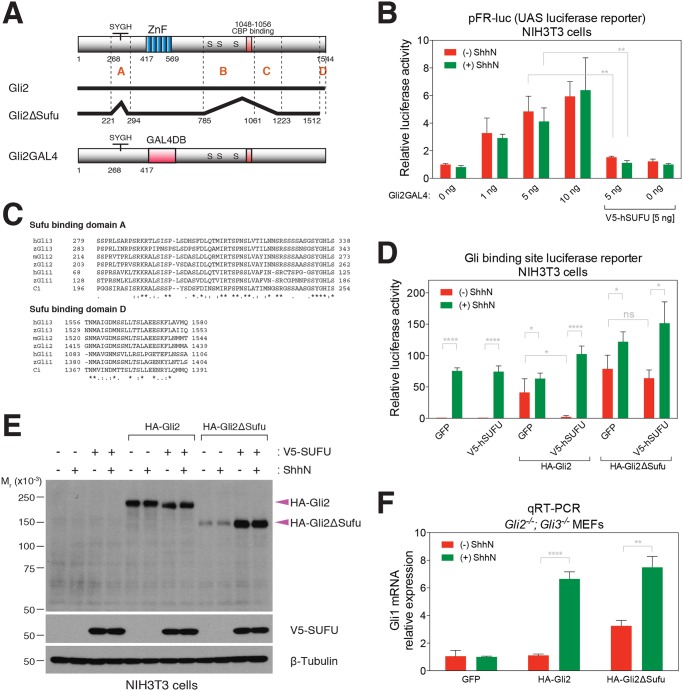
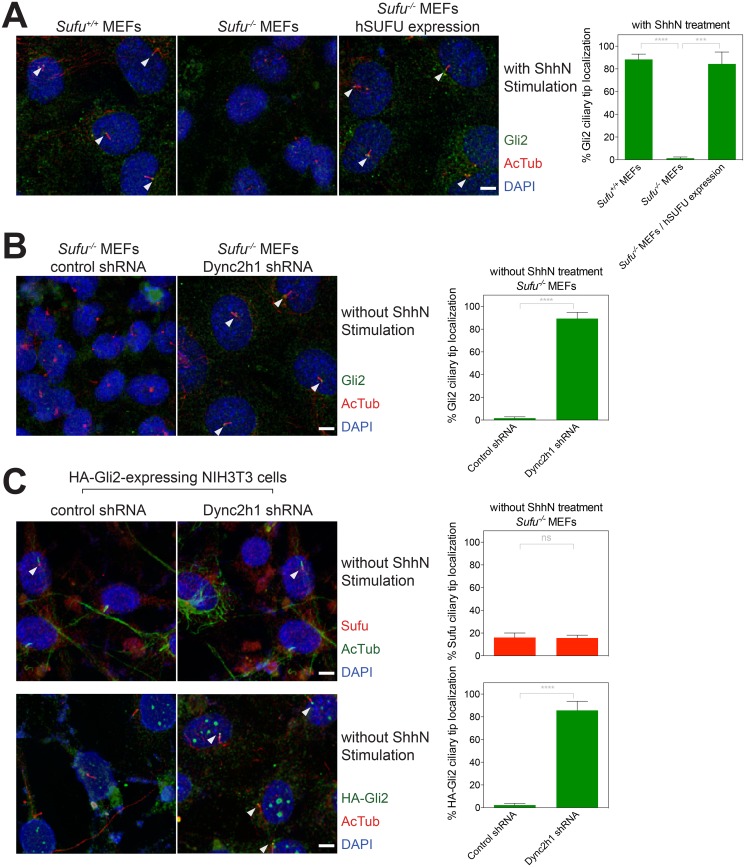
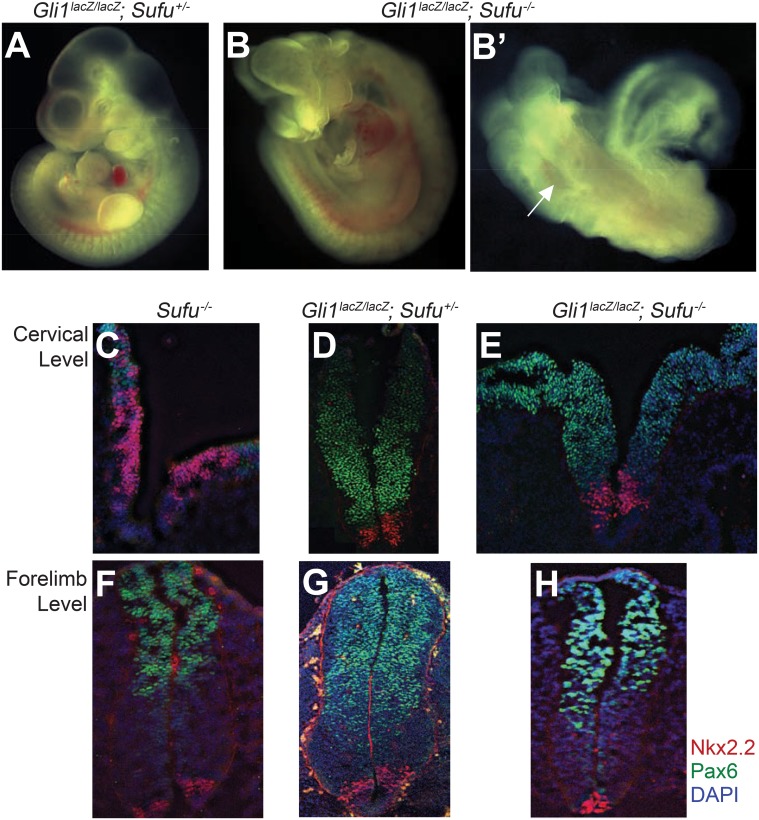
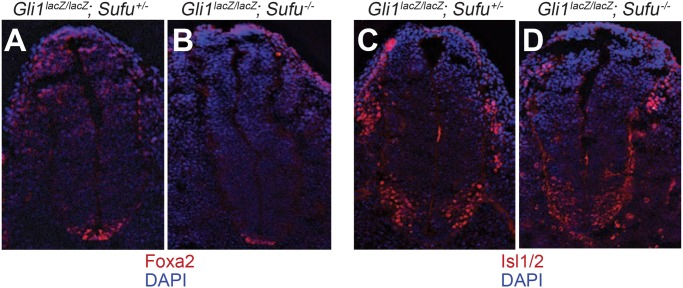
Suppressor of fused (Su(fu)/Sufu), one of the most conserved components of the Hedgehog (Hh) signaling pathway, binds Ci/Gli transcription factors and impedes activation of target gene expression. In Drosophila, the Su(fu) mutation has a minimal phenotype, and we show here that Ci transcriptional activity in large part is regulated independently of Su(fu) by other pathway components. Mutant mice lacking Sufu in contrast show excessive pathway activity and die as embryos with patterning defects. Here we show that in cultured cells Hh stimulation can augment transcriptional activity of a Gli2 variant lacking Sufu interaction and, surprisingly, that regulation of Hh pathway targets is nearly normal in the neural tube of Sufu-/- mutant embryos that also lack Gli1 function. Some degree of Hh-induced transcriptional activation of Ci/Gli thus can occur independently of Sufu in both flies and mammals. We further note that Sufu loss can also reduce Hh induction of high-threshold neural tube fates, such as floor plate, suggesting a possible positive pathway role for Sufu.
Conflict of interest statement
Figures
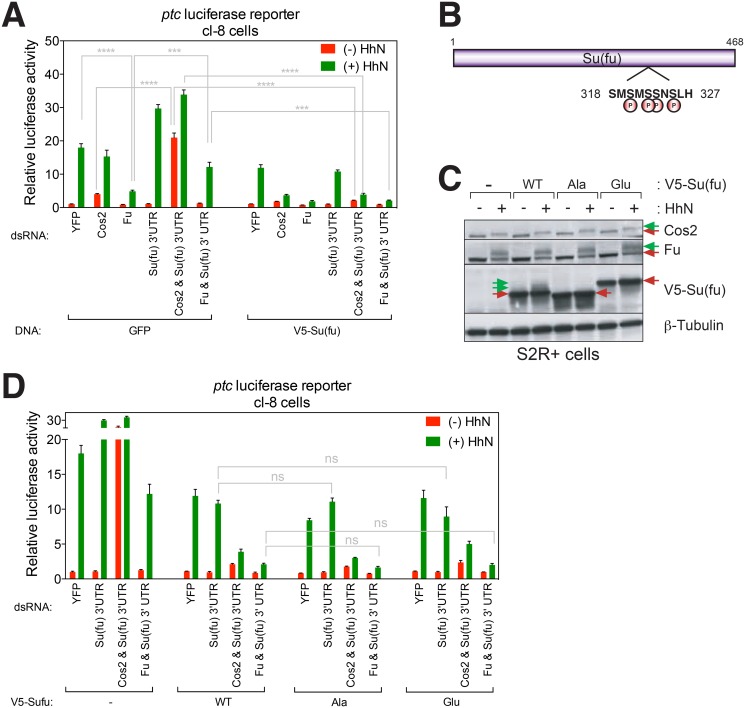
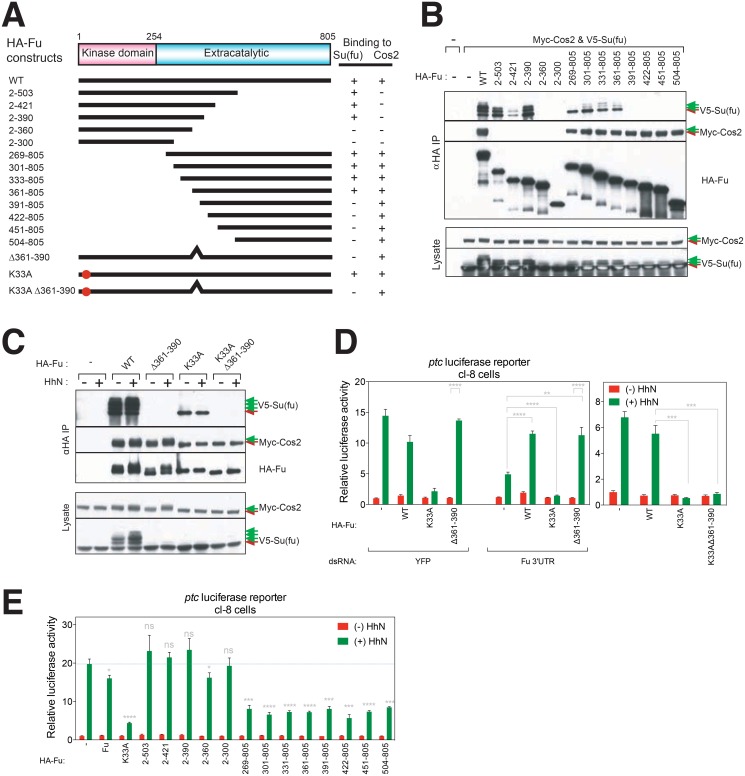
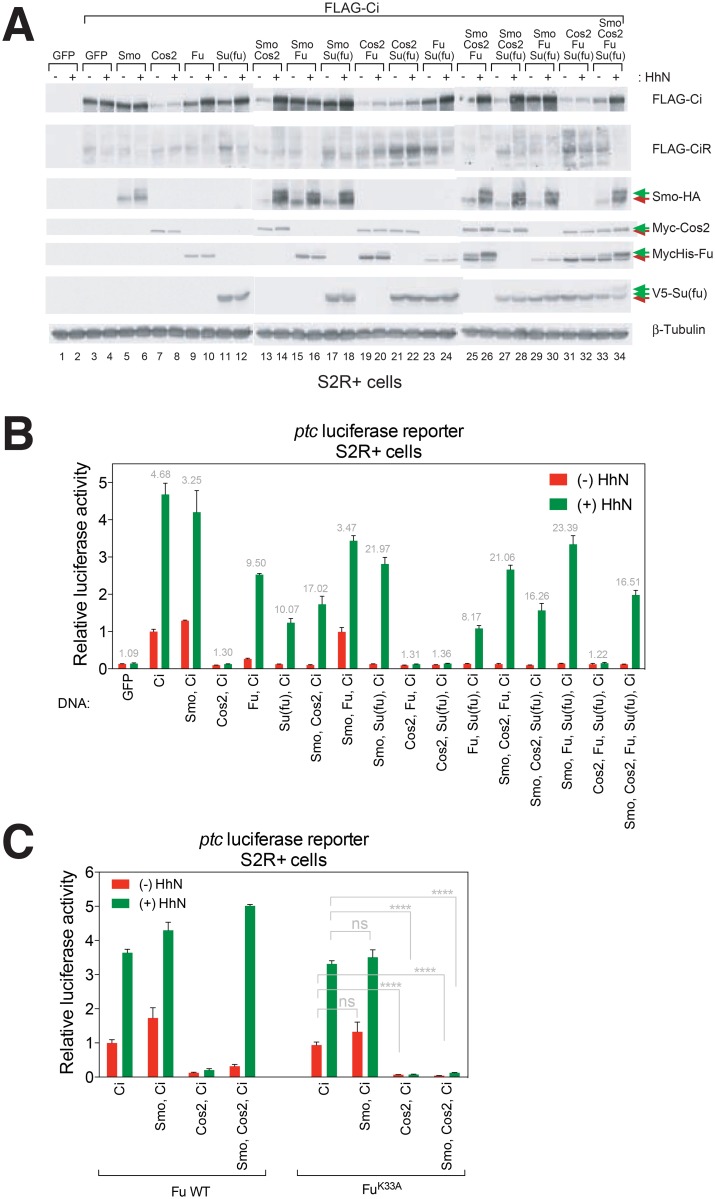
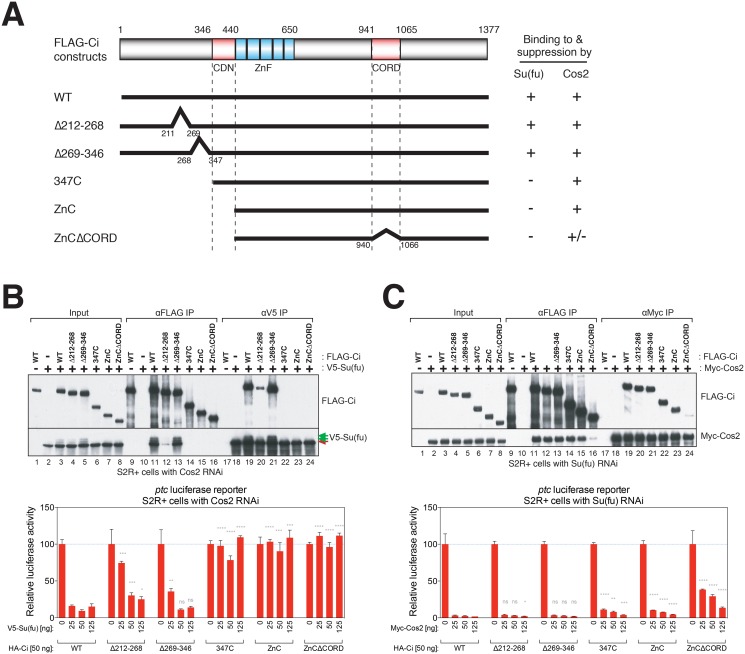
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
