

Absence of Vitamin K-Dependent γ-Carboxylation in Human Periostin Extracted from Fibrotic Lung or Secreted from a Cell Line Engineered to Optimize γ-Carboxylation
- PMID: 26273833
- PMCID: PMC4537219
- DOI: 10.1371/journal.pone.0135374
Absence of Vitamin K-Dependent γ-Carboxylation in Human Periostin Extracted from Fibrotic Lung or Secreted from a Cell Line Engineered to Optimize γ-Carboxylation
Abstract
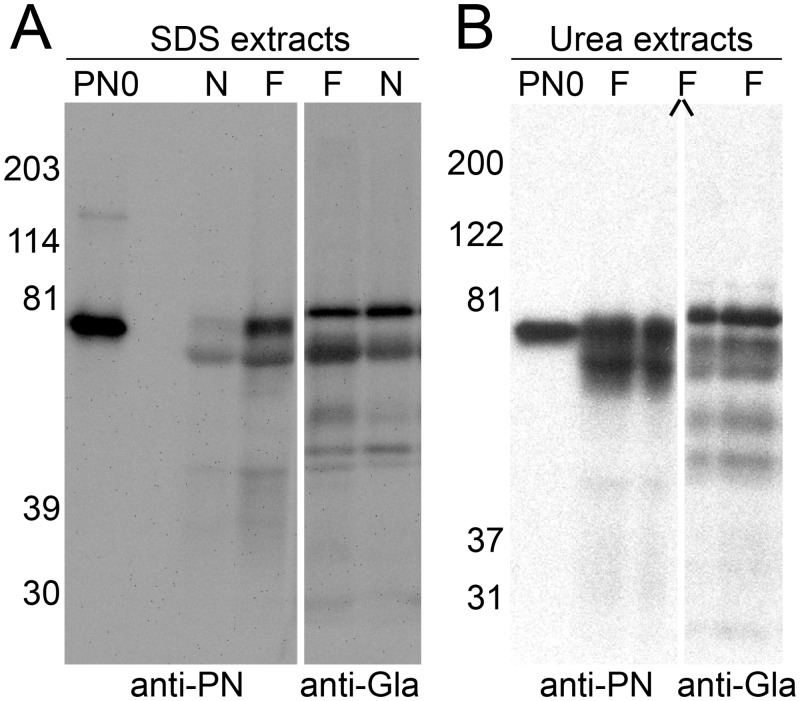
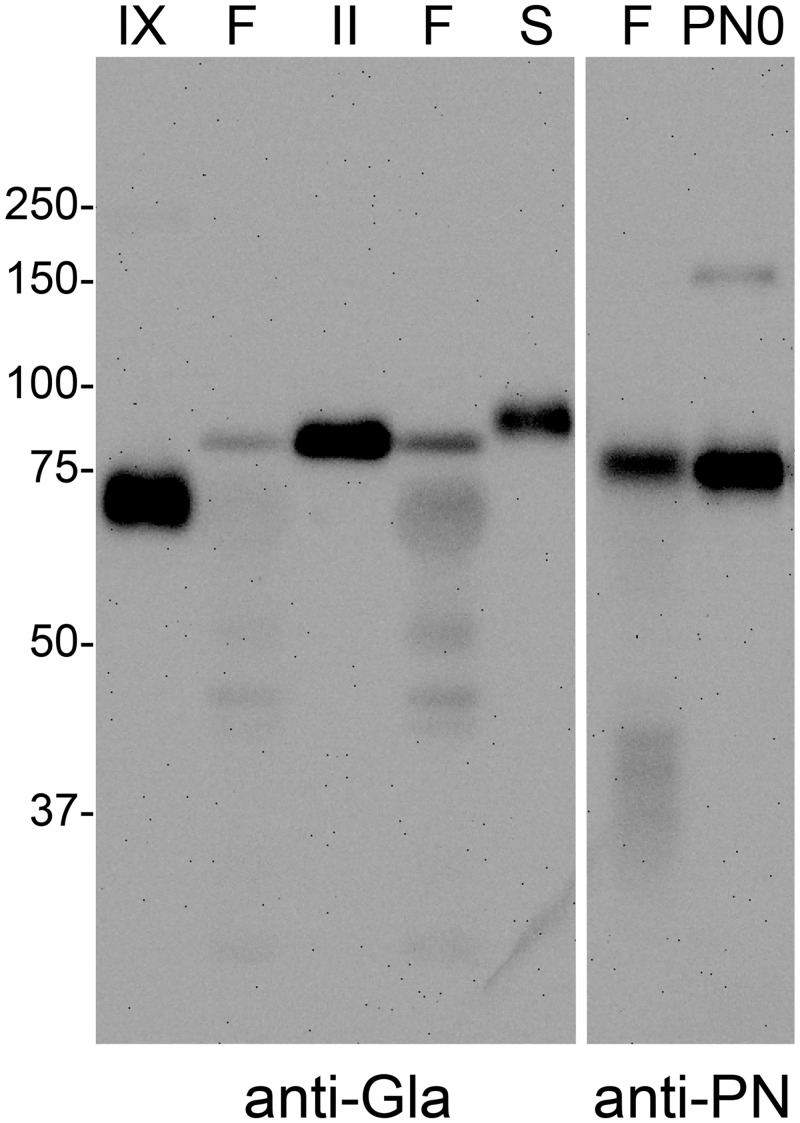
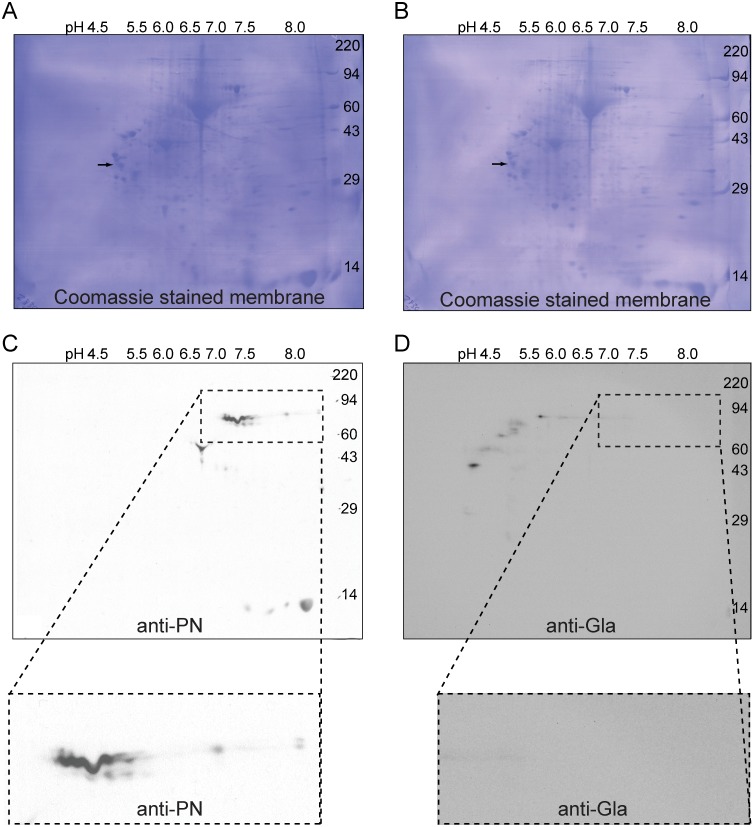
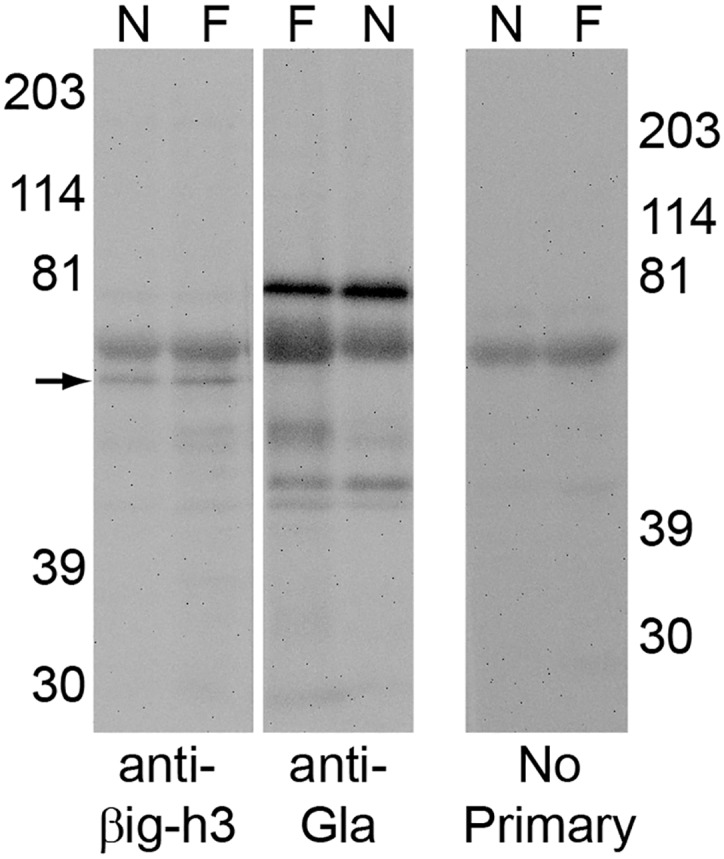
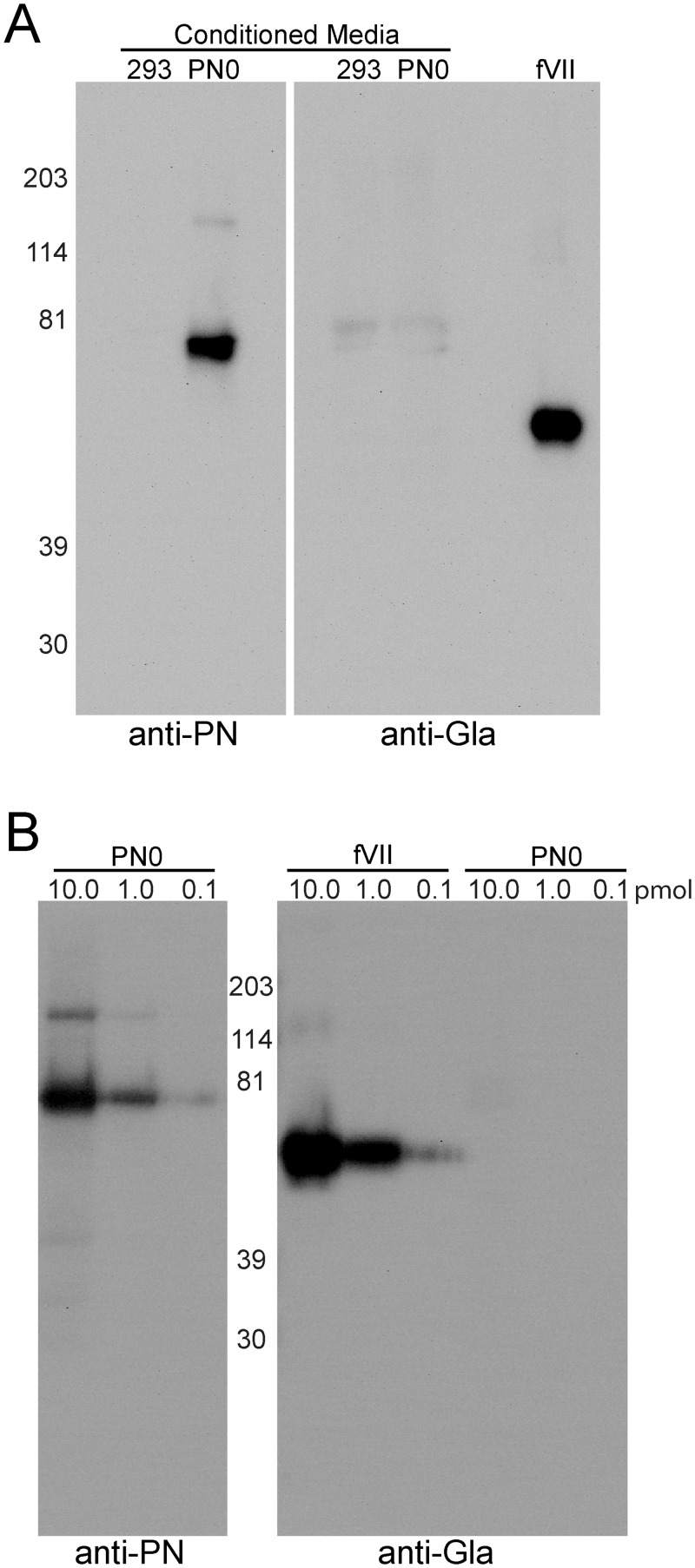
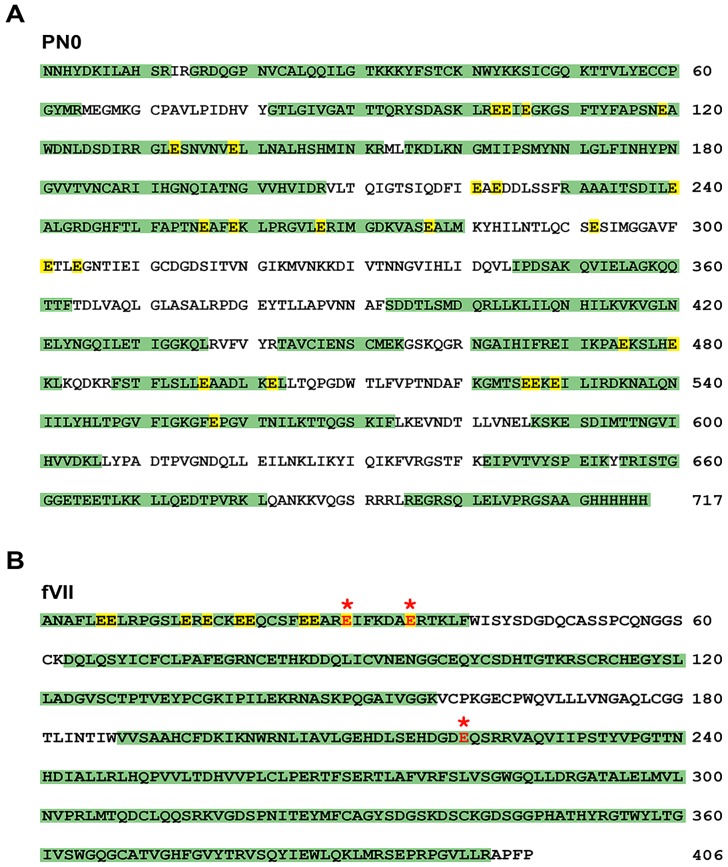
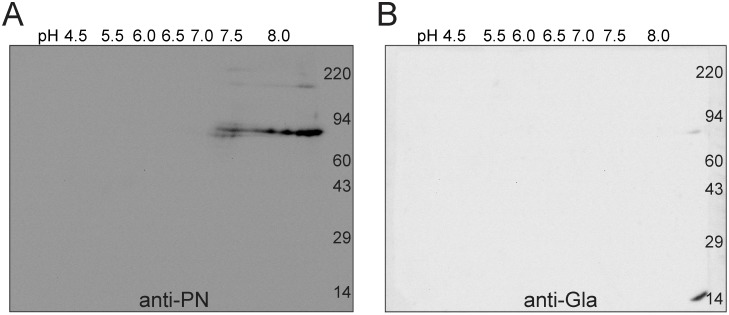
Periostin (PN, gene name POSTN) is an extracellular matrix protein that is up-regulated in bronchial epithelial cells and lung fibroblasts by TH-2 cytokines. Its paralog, TGF-β-induced protein (βig-h3, gene name TGFBI), is also expressed in the lung and up-regulated in bronchial myofibroblasts by TGF-β. PN and βig-h3 contain fasciclin 1 modules that harbor putative recognition sequences for γ-glutamyl carboxylase and are annotated in UniProt as undergoing vitamin K-dependent γ-carboxylation of multiple glutamic acid residues. γ-carboxylation profoundly alters activities of other proteins subject to the modification, e.g., blood coagulation factors, and would be expected to alter the structure and function of PN and βig-h3. To analyze for the presence of γ-carboxylation, proteins extracted from fibrotic lung were reacted with monoclonal antibodies specific for PN, βig-h3, or modification with γ-carboxyglutamic acid (Gla). In Western blots of 1-dimensional gels, bands stained with anti-PN or -βig-h3 did not match those stained with anti-Gla. In 2-dimensional gels, anti-PN-positive spots had pIs of 7.0 to >8, as expected for the unmodified protein, and there was no overlap between anti-PN-positive and anti-Gla-positive spots. Recombinant PN and blood coagulation factor VII were produced in HEK293 cells that had been transfected with vitamin K 2, 3-epoxide reductase C1 to optimize γ-carboxylation. Recombinant PN secreted from these cells did not react with anti-Gla antibody and had pIs similar to that found in extracts of fibrotic lung whereas secreted factor VII reacted strongly with anti-Gla antibody. Over 67% coverage of recombinant PN was achieved by mass spectrometry, including peptides with 19 of the 24 glutamates considered targets of γ-carboxylation, but analysis revealed no modification. Over 86% sequence coverage and three modified glutamic acid residues were identified in recombinant fVII. These data indicate that PN and βig-h3 are not subject to vitamin K-dependent γ-carboxylation.
Conflict of interest statement
Figures
References
-
- Skonier J, Neubauer M, Madisen L, Bennett K, Plowman GD, Purchio AF. cDNA cloning and sequence analysis of beta ig-h3, a novel gene induced in a human adenocarcinoma cell line after treatment with transforming growth factor-beta. DNA and cell biology. 1992;11(7):511–22. . - PubMed
-
- Mann KG, Nesheim ME, Church WR, Haley P, Krishnaswamy S. Surface-dependent reactions of the vitamin K-dependent enzyme complexes. Blood. 1990;76(1):1–16. . - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
