

Melatonin biosynthesis requires N-acetylserotonin methyltransferase activity of caffeic acid O-methyltransferase in rice
- PMID: 26276868
- PMCID: PMC4623696
- DOI: 10.1093/jxb/erv396
Melatonin biosynthesis requires N-acetylserotonin methyltransferase activity of caffeic acid O-methyltransferase in rice
Abstract
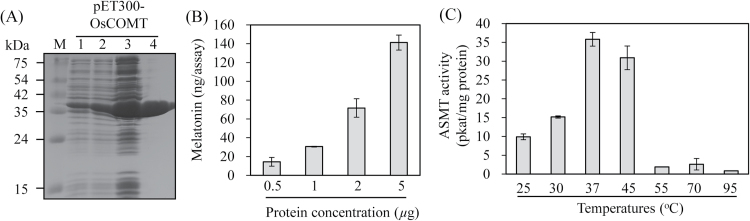
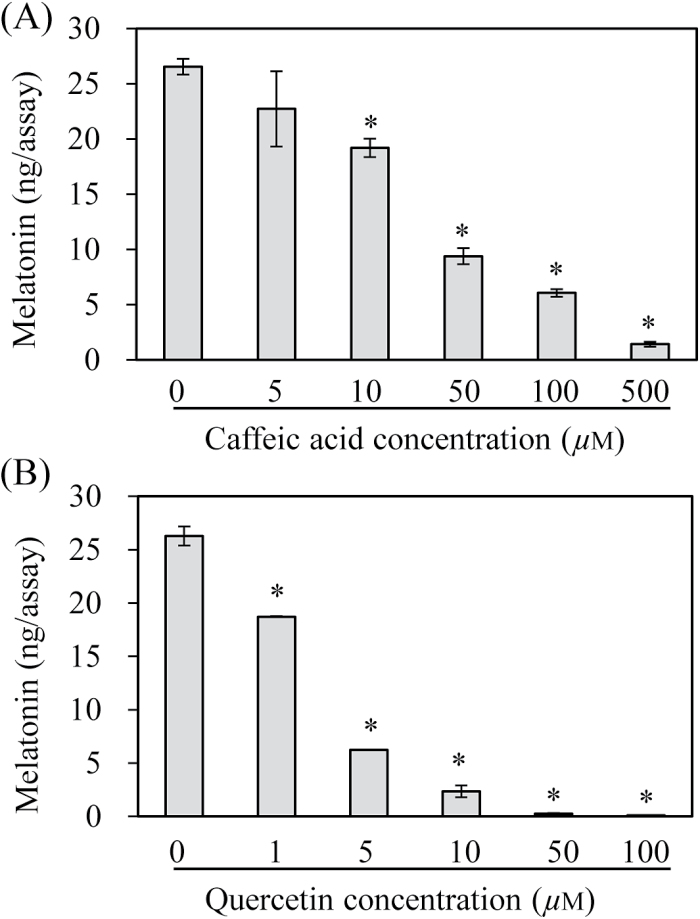
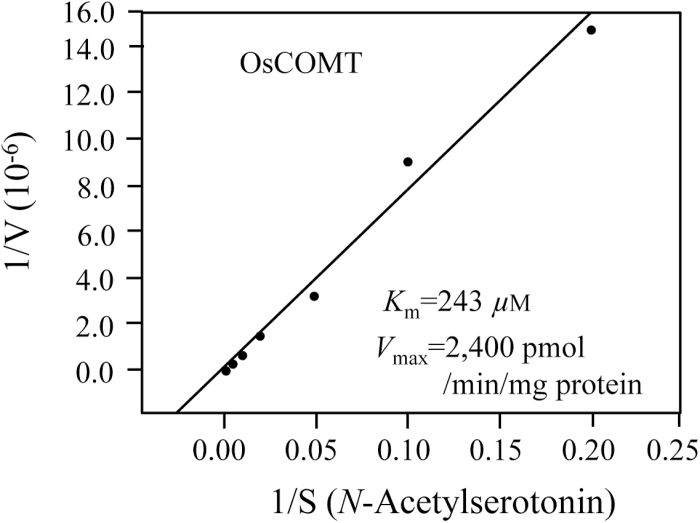
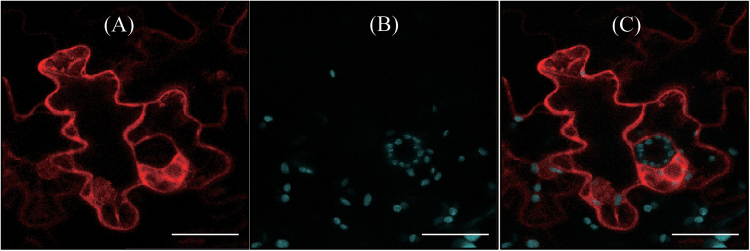
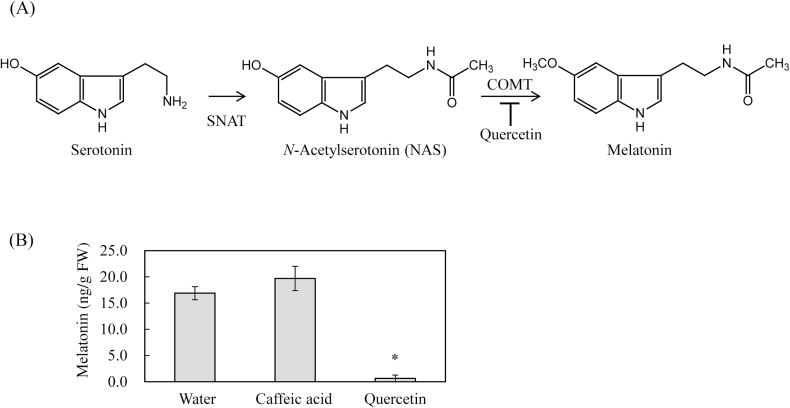
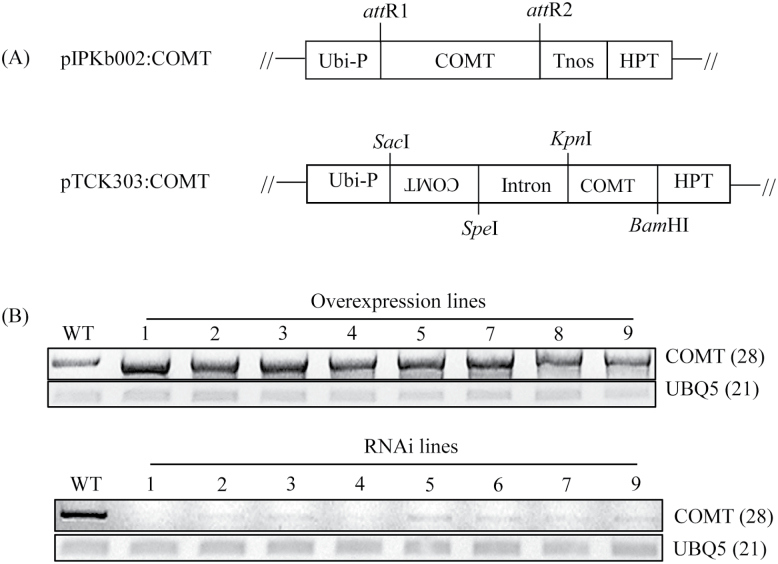
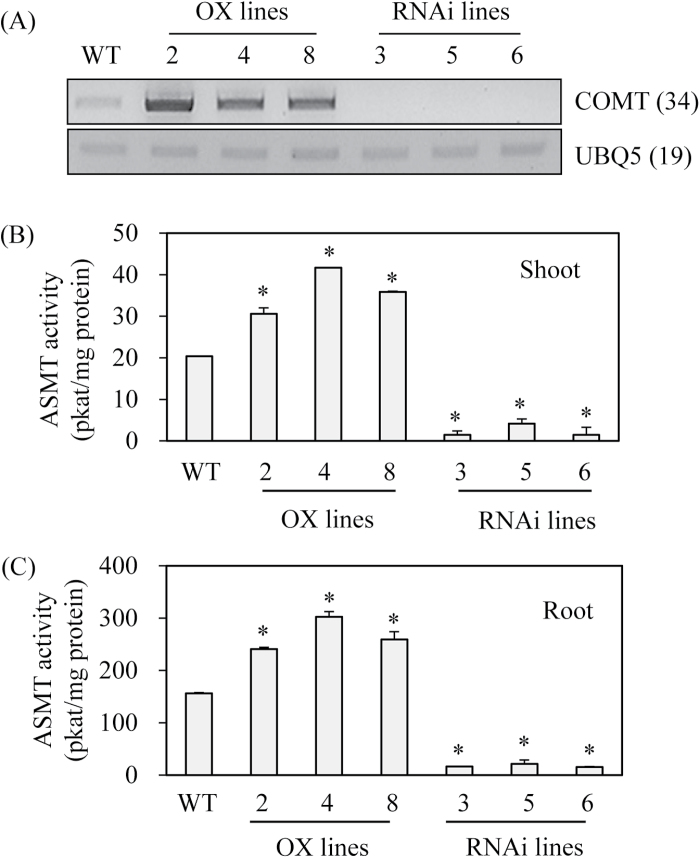
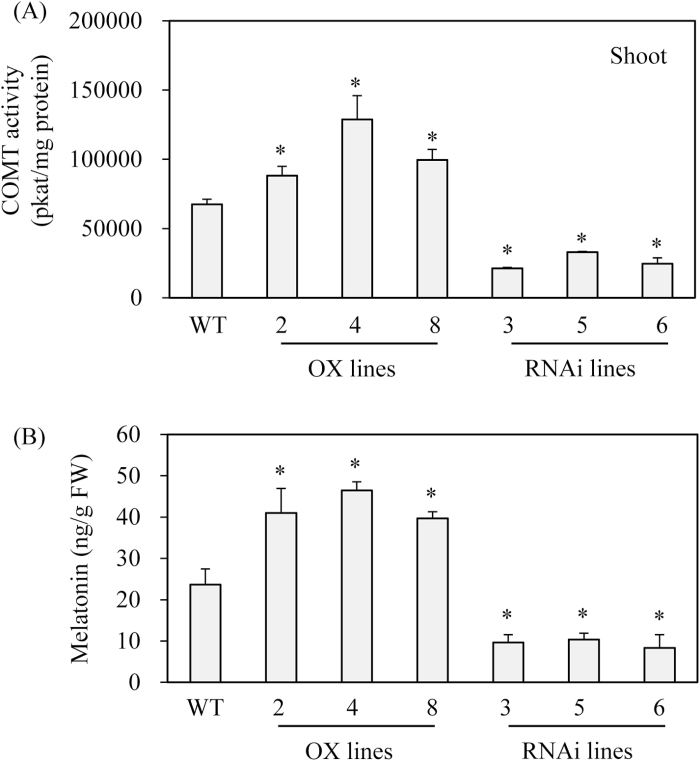
Caffeic acid O-methyltransferase (COMT) methylates N-acetylserotonin into melatonin; that is, it has N-acetylserotonin O-methyltransferase (ASMT) activity. The ASMT activity of COMT was first detected in Arabidopsis thaliana COMT (AtCOMT). To confirm the involvement of COMT on melatonin synthesis in other plant species, the ASMT activity of a COMT from rice (Oryza sativa) (OsCOMT) was evaluated. Purified recombinant OsCOMT protein from Escherichia coli was used to validate the high ASMT activity of OsCOMT, similar to that of AtCOMT. The K m and V max values for the ASMT activity of OsCOMT were 243 µM and 2400 pmol min(-1) mg protein(-1), which were similar to those of AtCOMT. Similar to AtCOMT, OsCOMT was localized in the cytoplasm. In vitro ASMT activity was significantly inhibited by either caffeic acid or quercetin in a dose-dependent manner. Analogously, in vivo production of melatonin was significantly inhibited by quercetin in 4-week-old detached rice leaves. Lastly, the transgenic rice plants overexpressing rice COMT showed an increase in melatonin levels whereas transgenic rice plants suppressing the rice COMT had a significant decrease on melatonin levels, suggestive of the direct role of COMT in melatonin biosynthesis in plants.
Keywords: N-Acetylserotonin O-methyltransferase; caffeic acid O-methyltransferase; melatonin; transgenic rice..
© The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures

References
-
- Arnao MB. 2014. Phytomelatonin: Discovery, content, and role in plants. Advances in Botany e815769.
-
- Arnao MB, Hernández-Ruiz J. 2014. Melatonin: plant growth regulator and/or biostimulator during stress? Trends in Plant Science 19, 789–797. - PubMed
-
- Buchanan BB, Gruissem W, Jones RL. 2000. Biochemistry and molecular biology of plants . American Society of Plant Physiologists, Rockville, MD.
-
- Byeon Y, Lee HY, Lee K, Back K. 2014. a Caffeic acid O-methyltransferase is involved in the synthesis of melatonin by methylating N-acetylserotonin in Arabidopsis . Journal of Pineal Research 57, 219–227. - PubMed
-
- Byeon Y, Lee HY, Lee K, Park S, Back K. 2014. b Cellular localization and kinetics of the rice melatonin biosynthetic enzymes SNAT and ASMT. Journal of Pineal Research 56, 107–114. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
