

Unoprostone activation of BK (KCa1.1) channel splice variants
- PMID: 26277265
- PMCID: PMC4598293
- DOI: 10.1016/j.bbamem.2015.08.005
Unoprostone activation of BK (KCa1.1) channel splice variants
Abstract
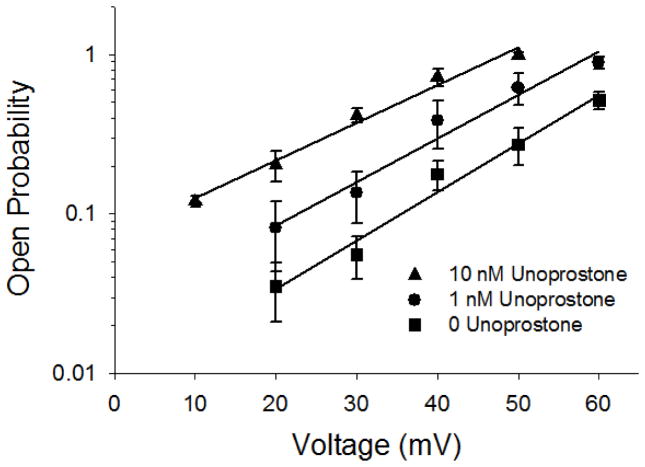
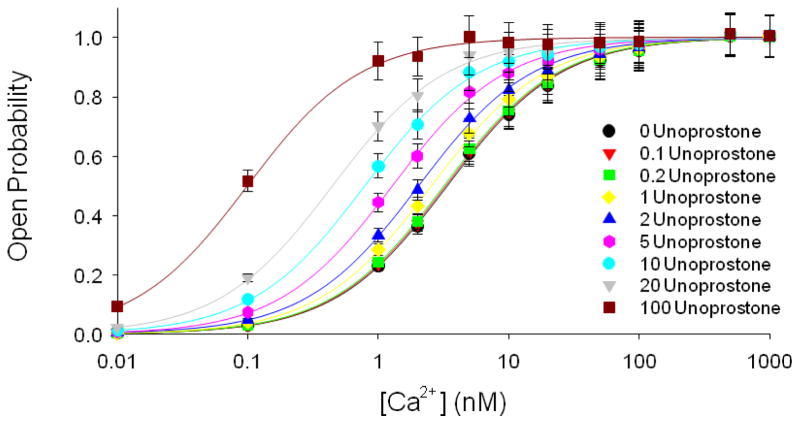
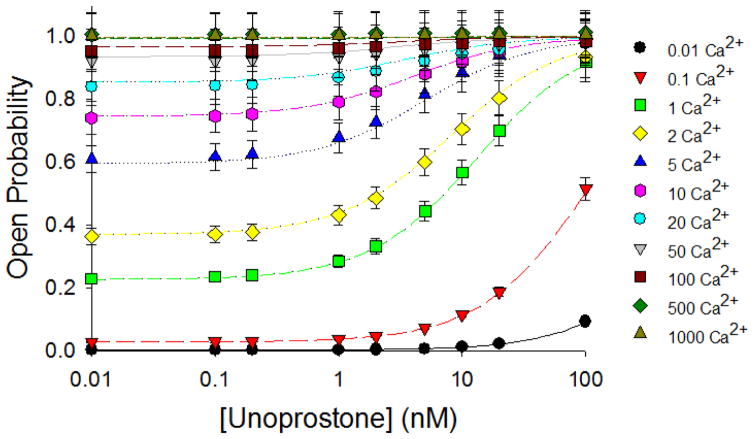
This investigation was conducted to study the relationship between intracellular Ca(2+) and activation of large conductance Ca(2+)-activated K(+) (BK) currents by unoprostone, the first synthetic docosanoid. We used HEK293 cells stably transfected with two BK channel splice variants, one sensitive to unoprostone and the other insensitive. We examined the effects of unoprostone on channel activity in excised inside-out patches and cell-attached patches. The half-maximal stimulation of the sensitive BK channels by Ca(2+) was shifted from 3.4±0.017 nM to 0.81±.0058 nM in the presence of 10 nM unoprostone. There was no effect on insensitive channels even at unoprostone concentrations as high as 1000 nM. There was no effect of unoprostone on the voltage dependence of the BK channels. Changes in open probability and effects of Ca(2+) and unoprostone were best described by a synergistic binding model. These data would suggest that Ca(2+) and unoprostone were binding to sites close to one another on the channel protein and that unoprostone binding causes the affinity of the calcium binding site to increase. This idea is consistent with three dimensional models of the Ca(2+) binding site and a putative unoprostone binding domain. Our results have important implications for the clinical use of unoprostone to activate BK channels. Channel activation will be limited if intracellular Ca(2+) is not elevated.
Keywords: BK channels; Ca(2+)-dependence; KCNMA1; Rescula®; Single channels; Unoprostone.
Copyright © 2015 Elsevier B.V. All rights reserved.
Figures







Similar articles
-
Unoprostone isopropyl and metabolite M1 activate BK channels and prevent ET-1-induced [Ca²⁺]i increases in human trabecular meshwork and smooth muscle.Invest Ophthalmol Vis Sci. 2012 Aug 7;53(9):5178-89. doi: 10.1167/iovs.11-9046. Invest Ophthalmol Vis Sci. 2012. PMID: 22786902
-
Cellular and molecular effects of unoprostone as a BK channel activator.Biochim Biophys Acta. 2007 May;1768(5):1083-92. doi: 10.1016/j.bbamem.2006.12.015. Epub 2007 Jan 3. Biochim Biophys Acta. 2007. PMID: 17307133
-
Functional validation of Ca2+-binding residues from the crystal structure of the BK ion channel.Biochim Biophys Acta Biomembr. 2018 Apr;1860(4):943-952. doi: 10.1016/j.bbamem.2017.09.023. Epub 2017 Sep 29. Biochim Biophys Acta Biomembr. 2018. PMID: 28966112
-
KCNMA1-linked channelopathy.J Gen Physiol. 2019 Oct 7;151(10):1173-1189. doi: 10.1085/jgp.201912457. Epub 2019 Aug 19. J Gen Physiol. 2019. PMID: 31427379 Free PMC article. Review.
-
BK channel activators and their therapeutic perspectives.Front Physiol. 2014 Oct 9;5:389. doi: 10.3389/fphys.2014.00389. eCollection 2014. Front Physiol. 2014. PMID: 25346695 Free PMC article. Review.
Cited by
-
BK Channels in the Central Nervous System.Int Rev Neurobiol. 2016;128:281-342. doi: 10.1016/bs.irn.2016.04.001. Epub 2016 May 13. Int Rev Neurobiol. 2016. PMID: 27238267 Free PMC article. Review.
References
-
- Abelson MB, McLaughlin J. A Look at Typical Non-Responders to Drug Therapy and the Factors That Have Made Them That Way. 2011
-
- Bers DM, Patton CW, Nuccitelli R. A practical guide to the preparation of Ca(2+) buffers. Methods Cell Biol. 2010;99:1–26. - PubMed
-
- Blumenfeld A, Gennings C, Cady R. Pharmacological synergy: the next frontier on therapeutic advancement for migraine. Headache. 2012;52:636–647. - PubMed
-
- Boland MV, Ervin AM, Friedman D, Jampel H, Hawkins B, Volenweider D, Chelladurai Y, Ward D, Suarez-Cuervo C, Robinson KA. 2012 - PubMed
-
- Clark AF, Lane D, Wilson K, Miggans ST, McCartney MD. Inhibition of dexamethasone-induced cytoskeletal changes in cultured human trabecular meshwork cells by tetrahydrocortisol. Invest Ophthalmol Vis Sci. 1996;37:805–813. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
