

CARF (Collaborator of ARF) overexpression in p53-deficient cells promotes carcinogenesis
- PMID: 26278998
- PMCID: PMC5528716
- DOI: 10.1016/j.molonc.2015.07.003
CARF (Collaborator of ARF) overexpression in p53-deficient cells promotes carcinogenesis
Abstract
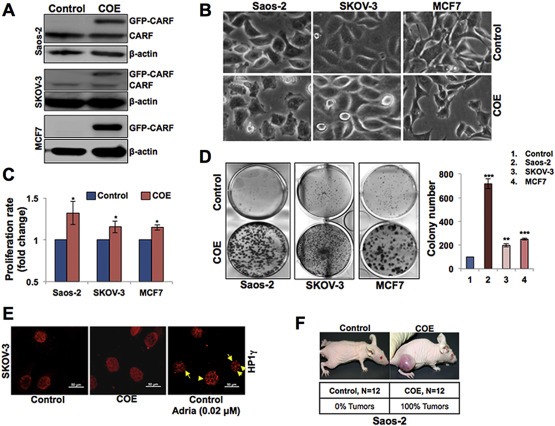
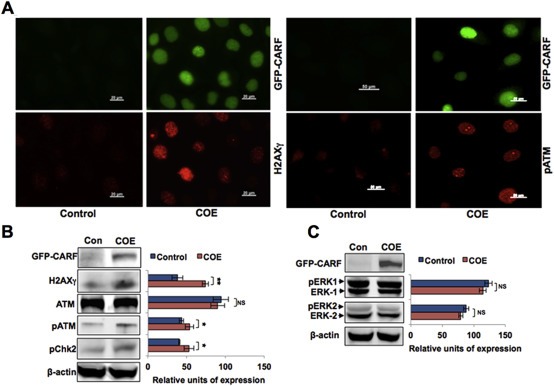
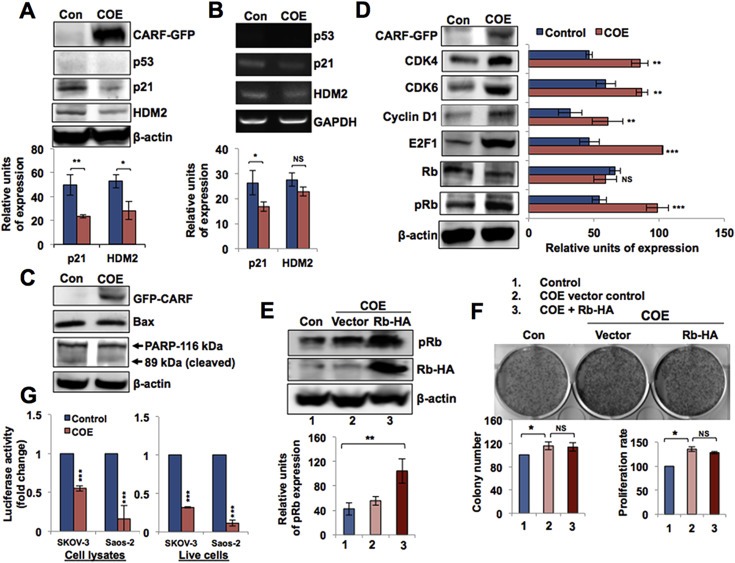
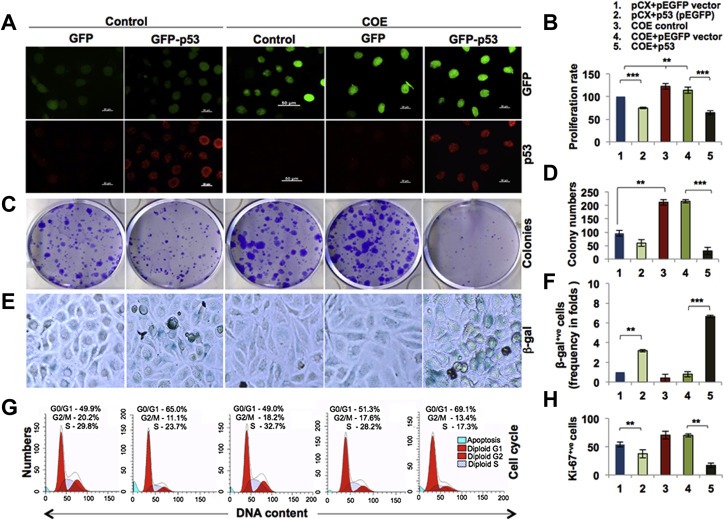
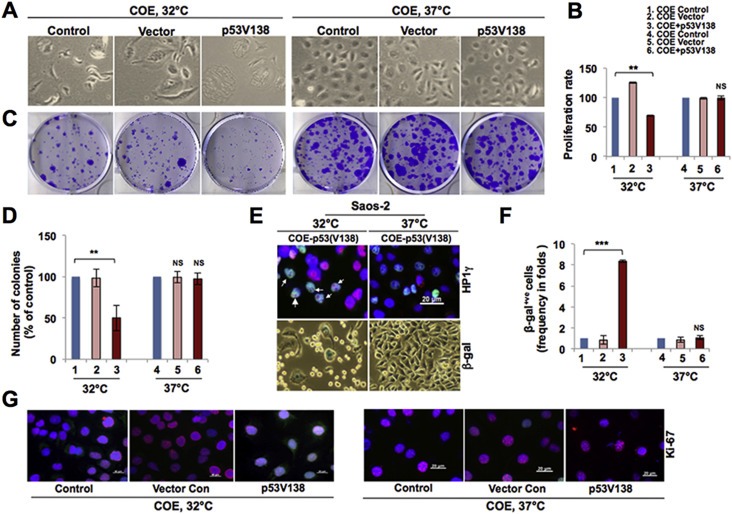
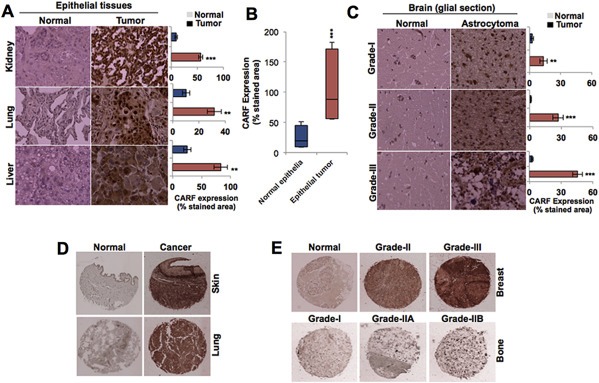
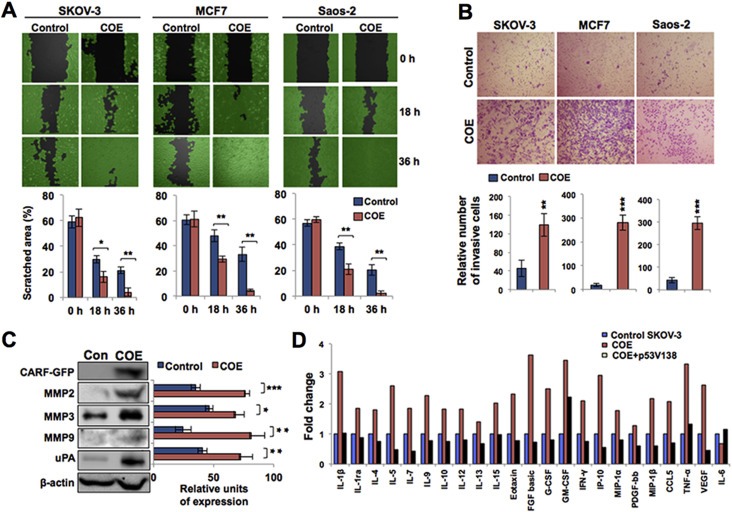
Collaborator of ARF (CARF), initially identified as a binding partner of ARF (Alternate Reading Frame), has been shown to activate ARF-p53 pathway by multiple ways including stabilization of ARF and p53 tumor suppressor proteins, and transcriptional repression of a p53 antagonist, HDM2. Level of CARF expression was shown to determine fate of cells. Whereas its knockdown caused apoptosis, its over- and super-expressions caused senescence and increase in malignant properties of cancer cells, respectively, and were closely linked to increase and decrease in p53 activity. Using p53-compromised cancer cells, we demonstrate that CARF induces growth arrest when wild type p53 is present and in p53-absence, it promotes carcinogenesis. Biochemical analyses on CARF-induced molecular signaling revealed that in p53-null cells, it caused transcriptional repression of p21(WAF1) leading to increase in CDK4, CDK6, pRb and E2F1 resulting in continued cell cycle progression. Furthermore, it instigated increase in migration and invasion of cancer cells that was marked by upregulation of MMP2, MMP3, MMP9, uPA, several interleukins and VEGF expression. Consistent with these findings, we found that human clinical samples of epithelial and glial cancers (frequently marked by loss of p53 function) possessed high level of CARF expression showing a relationship with cancer aggressiveness. The data demonstrated that CARF could be considered as a diagnostic marker and a therapeutic target in p53-compromised malignancies.
Keywords: CARF (Collaborator of ARF); Growth arrest; Overexpression; Proliferation; p53 reconstitution; p53-Deficient cancer cells.
Copyright © 2015 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Alla, V. , Engelmann, D. , Niemetz, A. , Pahnke, J. , Schmidt, A. , Kunz, M. , Emmrich, S. , Steder, M. , Koczan, D. , Putzer, B.M. , 2010. E2F1 in melanoma progression and metastasis. J. Natl. Cancer Inst. 102, 127–133. - PubMed
-
- Bartek, J. , Bartkova, J. , Lukas, J. , 2007. DNA damage signalling guards against activated oncogenes and tumour progression. Oncogene. 26, 7773–7779. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
