

Bile Acids Trigger GLP-1 Release Predominantly by Accessing Basolaterally Located G Protein-Coupled Bile Acid Receptors
- PMID: 26280129
- PMCID: PMC4606749
- DOI: 10.1210/en.2015-1321
Bile Acids Trigger GLP-1 Release Predominantly by Accessing Basolaterally Located G Protein-Coupled Bile Acid Receptors
Abstract
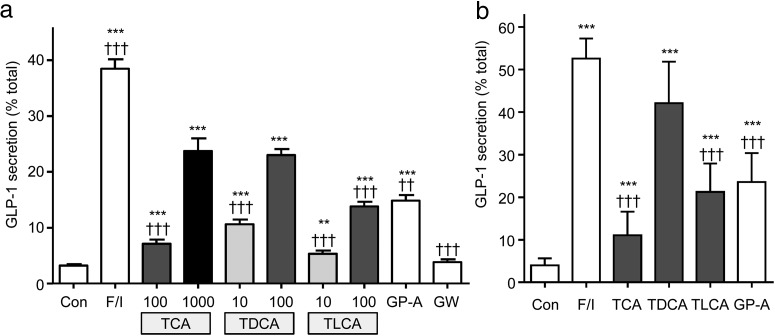
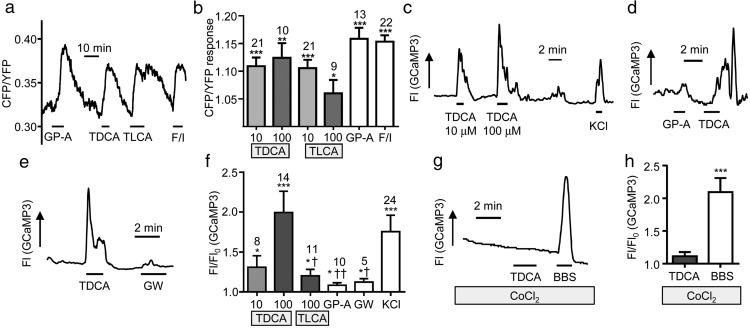
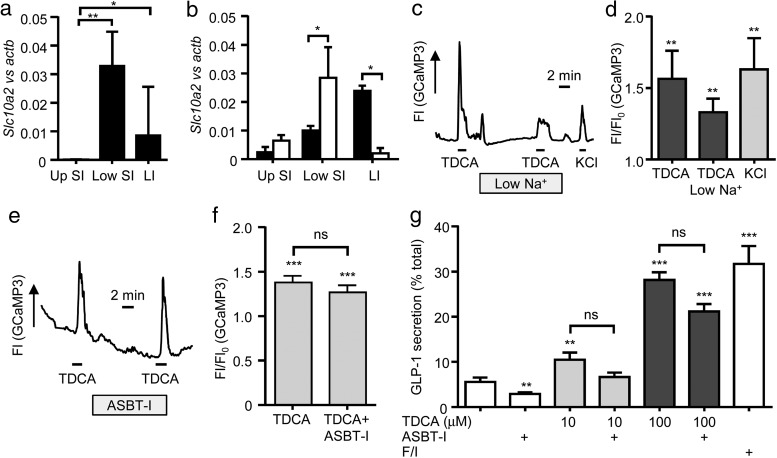
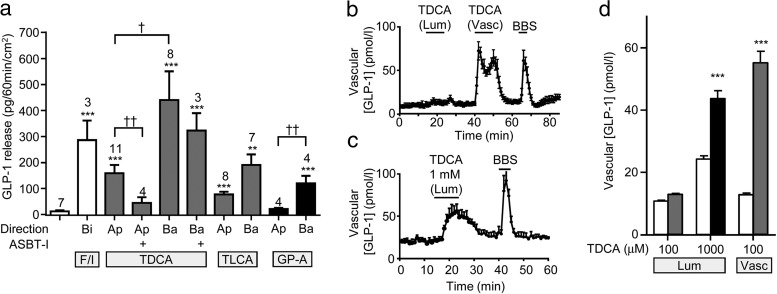
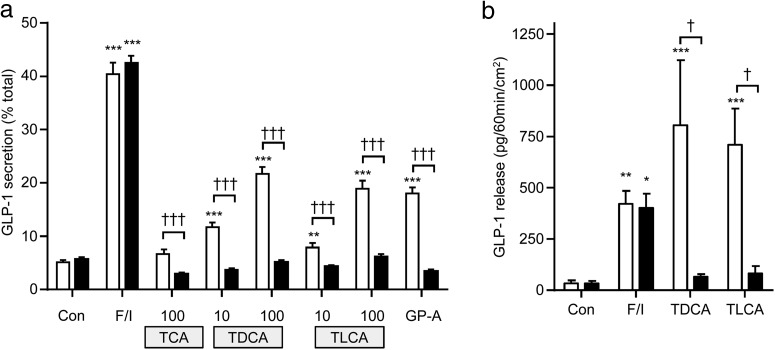
Bile acids are well-recognized stimuli of glucagon-like peptide-1 (GLP-1) secretion. This action has been attributed to activation of the G protein-coupled bile acid receptor GPBAR1 (TGR5), although other potential bile acid sensors include the nuclear farnesoid receptor and the apical sodium-coupled bile acid transporter ASBT. The aim of this study was to identify pathways important for GLP-1 release and to determine whether bile acids target their receptors on GLP-1-secreting L-cells from the apical or basolateral compartment. Using transgenic mice expressing fluorescent sensors specifically in L-cells, we observed that taurodeoxycholate (TDCA) and taurolithocholate (TLCA) increased intracellular cAMP and Ca(2+). In primary intestinal cultures, TDCA was a more potent GLP-1 secretagogue than taurocholate (TCA) and TLCA, correlating with a stronger Ca(2+) response to TDCA. Using small-volume Ussing chambers optimized for measuring GLP-1 secretion, we found that both a GPBAR1 agonist and TDCA stimulated GLP-1 release better when applied from the basolateral than from the luminal direction and that luminal TDCA was ineffective when intestinal tissue was pretreated with an ASBT inhibitor. ASBT inhibition had no significant effect in nonpolarized primary cultures. Studies in the perfused rat gut confirmed that vascularly administered TDCA was more effective than luminal TDCA. Intestinal primary cultures and Ussing chamber-mounted tissues from GPBAR1-knockout mice did not secrete GLP-1 in response to either TLCA or TDCA. We conclude that the action of bile acids on GLP-1 secretion is predominantly mediated by GPBAR1 located on the basolateral L-cell membrane, suggesting that stimulation of gut hormone secretion may include postabsorptive mechanisms.
Figures
References
-
- Baggio LL, Drucker DJ. Biology of incretins: GLP-1 and GIP. Gastroenterology. 2007;132:2131–2157. - PubMed
-
- Holst JJ. The physiology of glucagon-like peptide 1. Physiol Rev. 2007;87:1409–1439. - PubMed
-
- Tolhurst G, Reimann F, Gribble FM. Intestinal sensing of nutrients. Handb Exp Pharmacol. 2012;209:309–335. - PubMed
-
- Nauck MA. Incretin-based therapies for type 2 diabetes mellitus: properties, functions, and clinical implications. Am J Med. 2011;124(1 Suppl):S3–S18. - PubMed
-
- Jørgensen NB, Jacobsen SH, Dirksen C, et al. Acute and long-term effects of Roux-en-Y gastric bypass on glucose metabolism in subjects with type 2 diabetes and normal glucose tolerance. Am J Physiol Endocrinol Metab. 2012;303:E122–E131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
