

An atomic structure of human γ-secretase
- PMID: 26280335
- PMCID: PMC4568306
- DOI: 10.1038/nature14892
An atomic structure of human γ-secretase
Abstract
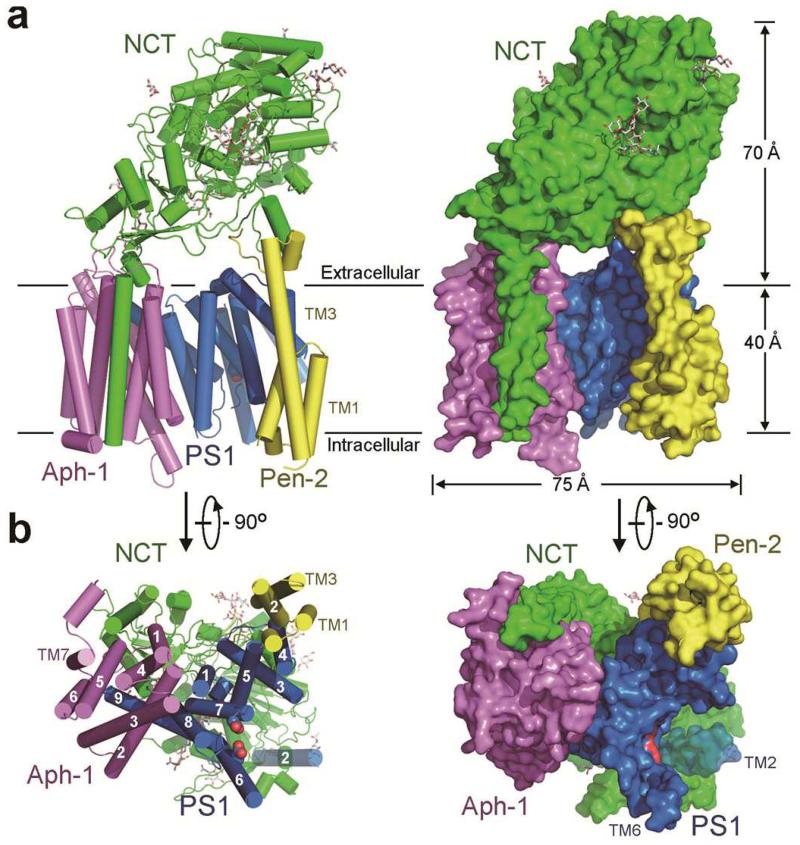
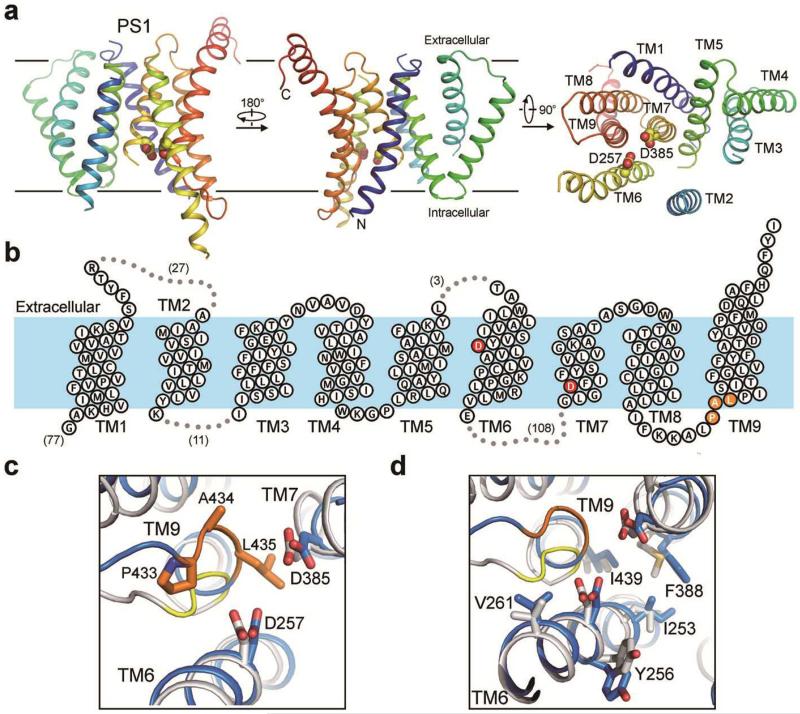
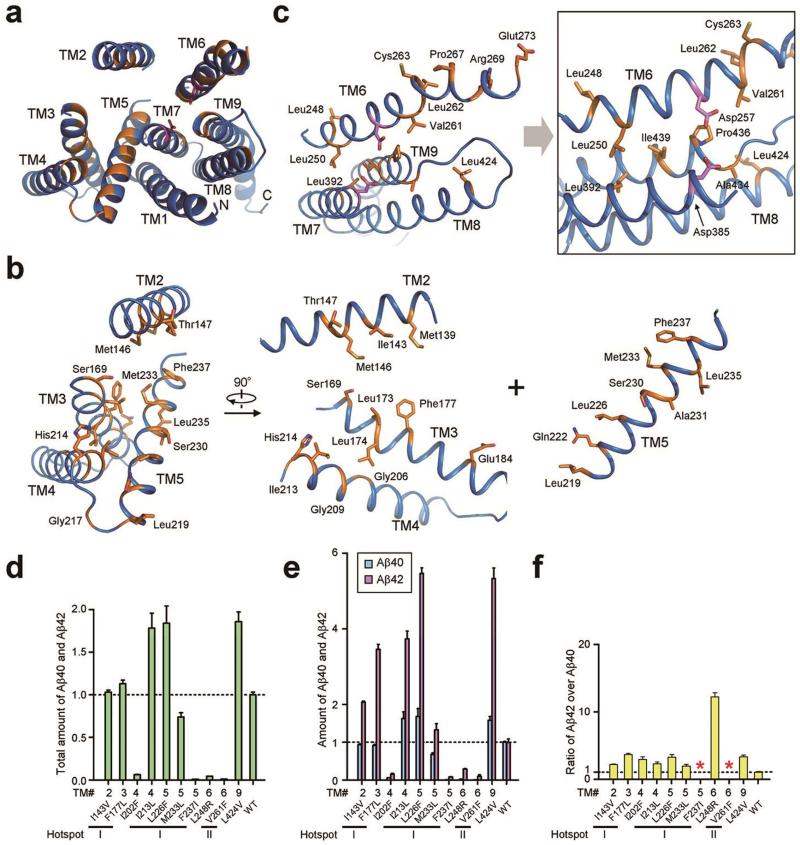
Dysfunction of the intramembrane protease γ-secretase is thought to cause Alzheimer's disease, with most mutations derived from Alzheimer's disease mapping to the catalytic subunit presenilin 1 (PS1). Here we report an atomic structure of human γ-secretase at 3.4 Å resolution, determined by single-particle cryo-electron microscopy. Mutations derived from Alzheimer's disease affect residues at two hotspots in PS1, each located at the centre of a distinct four transmembrane segment (TM) bundle. TM2 and, to a lesser extent, TM6 exhibit considerable flexibility, yielding a plastic active site and adaptable surrounding elements. The active site of PS1 is accessible from the convex side of the TM horseshoe, suggesting considerable conformational changes in nicastrin extracellular domain after substrate recruitment. Component protein APH-1 serves as a scaffold, anchoring the lone transmembrane helix from nicastrin and supporting the flexible conformation of PS1. Ordered phospholipids stabilize the complex inside the membrane. Our structure serves as a molecular basis for mechanistic understanding of γ-secretase function.
Figures










References
-
- Alzheimer A. About a peculiar disease of the cerebral cortex. Centralblatt für Nervenheilkunde Psychiatrie. 1907;30:177–179.
-
- Hardy JA, Higgins GA. Alzheimer’s disease: the amyloid cascade hypothesis. Science. 1992;256:184–5. - PubMed
-
- Goate A, Hardy J. Twenty years of Alzheimer’s disease-causing mutations. J Neurochem. 2012;120(Suppl 1):3–8. - PubMed
-
- Tanzi RE, Bertram L. Twenty years of the Alzheimer’s disease amyloid hypothesis: a genetic perspective. Cell. 2005;120:545–55. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
