

Targeted Disruption of the β2-Microglobulin Gene Minimizes the Immunogenicity of Human Embryonic Stem Cells
- PMID: 26285657
- PMCID: PMC4572902
- DOI: 10.5966/sctm.2015-0049
Targeted Disruption of the β2-Microglobulin Gene Minimizes the Immunogenicity of Human Embryonic Stem Cells
Abstract
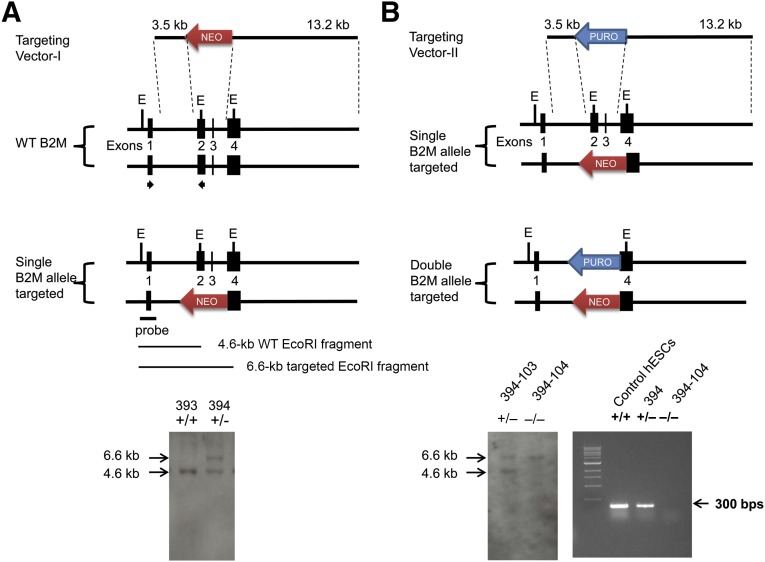
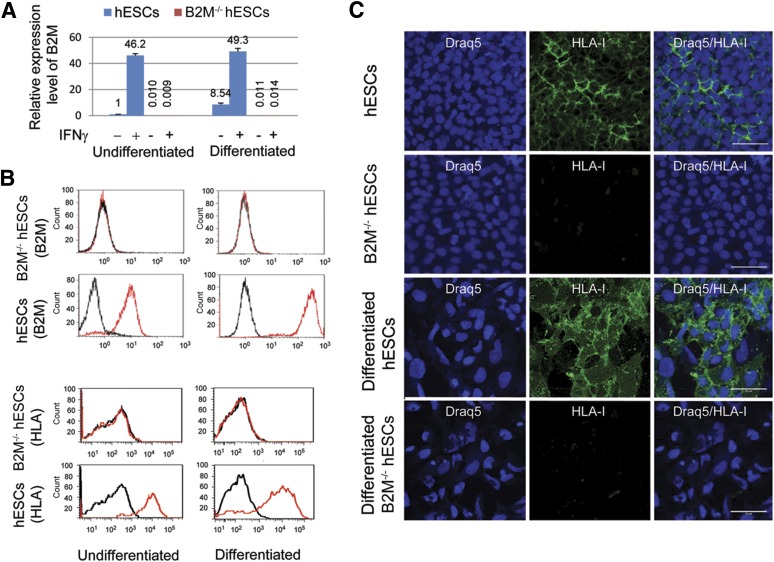
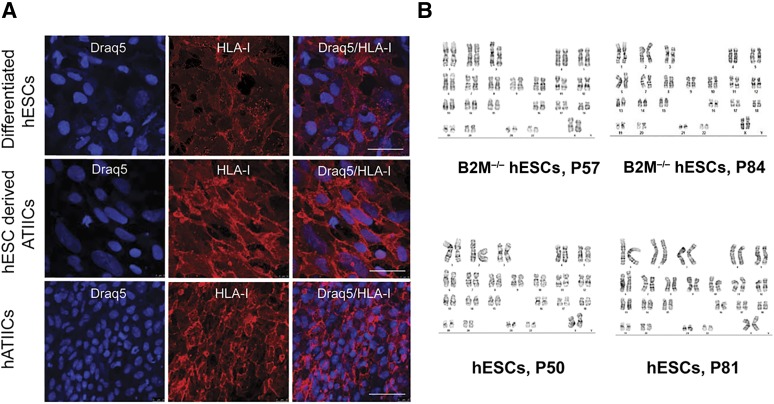
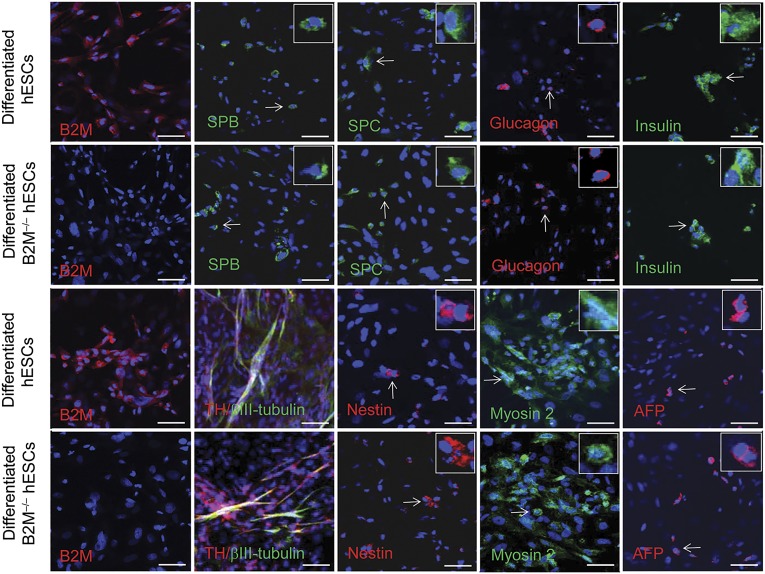
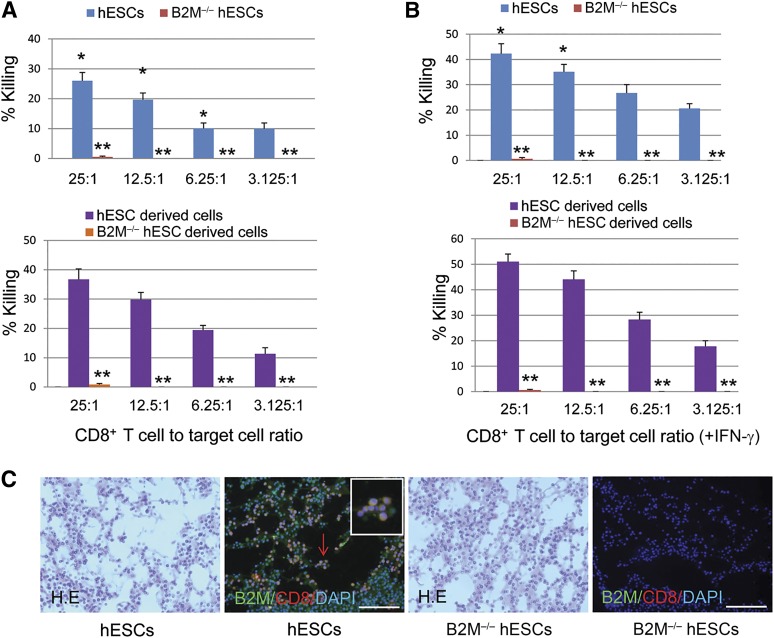
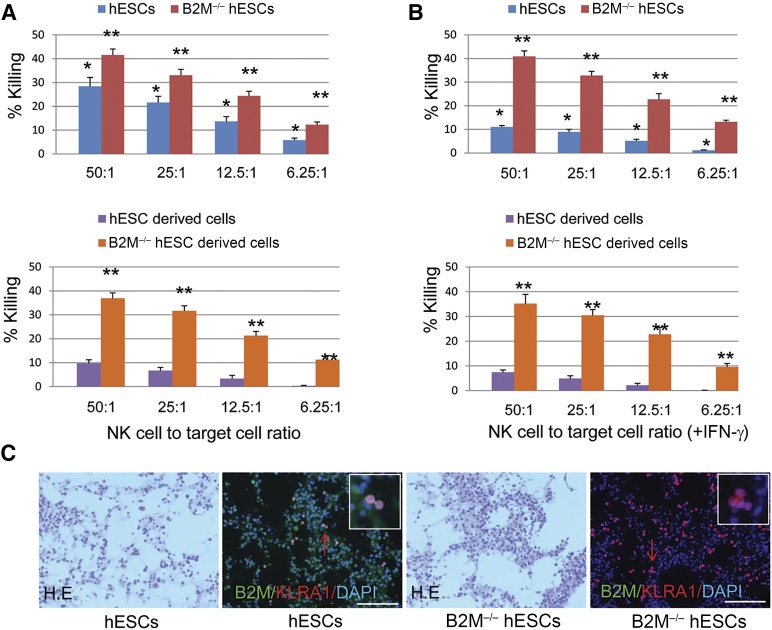
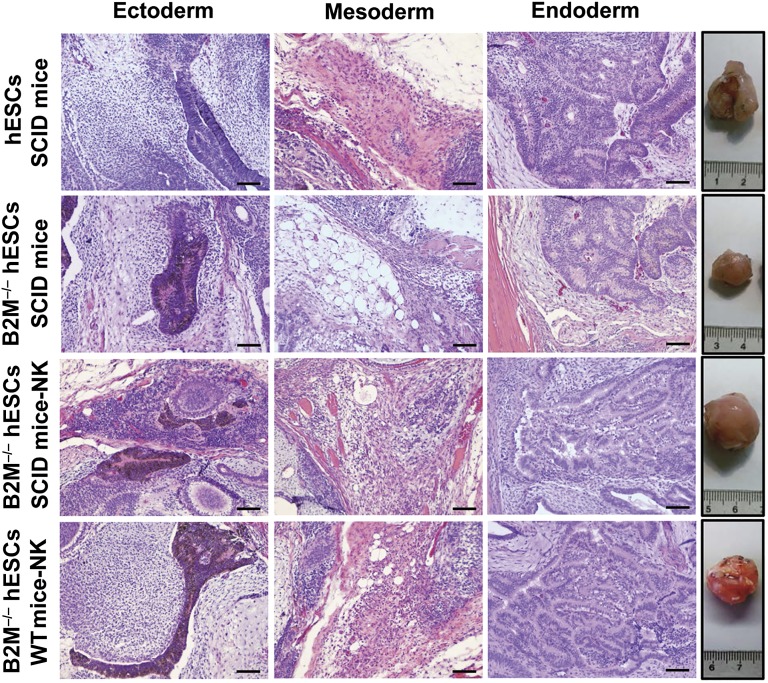
Human embryonic stem cells (hESCs) are a promising source of cells for tissue regeneration, yet histoincompatibility remains a major challenge to their clinical application. Because the human leukocyte antigen class I (HLA-I) molecules are the primary mediators of immune rejection, we hypothesized that cells derived from a hESC line lacking HLA-I expression could be transplanted without evoking a robust immune response from allogeneic recipients. In the present study, we used the replacement targeting strategy to delete exons 2 and 3 of β2-microglobulin on both gene alleles in hESCs. Because β2-microglobulin serves as the HLA-I light chain, disruption of the β2-microglobulin gene led to complete HLA-I deficiency on the cell surface of hESCs and their derivatives. Therefore, these cells were resistant to CD8+ T-cell-mediated destruction. Although interferon-γ (IFN-γ) treatment significantly induced β2-microglobulin expression, promoting CD8+ T cell-mediated killing of control hESCs and their derivatives, CD8+ T-cell-mediated cytotoxicity was barely observed with β2-microglobulin-null hESCs and their derivatives treated with IFN-γ. This genetic manipulation to disrupt HLA-I expression did not affect the self-renewal capacity, genomic stability, or pluripotency of hESCs. Despite being relatively sensitive to natural killer (NK) cell-mediated killing due to the lack of HLA-I expression, when transplanted into NK cell-depleted immunocompetent mice, β2-microglobulin-null hESCs developed into tumors resembling those derived from control hESCs in severe combined immunodeficiency mice. These results demonstrate that β2-microglobulin-null hESCs significantly reduce immunogenicity to CD8+ T cells and might provide a renewable source of cells for tissue regeneration without the need for HLA matching in the future.
Significance: This study reports the generation of a novel β2-microglobulin (B2M)-/- human embryonic stem cell (hESC) line. Differentiated mature cells from this line do not express cell surface human leukocyte antigen molecules even after interferon-γ stimulation and are resistant to alloreactive CD8+ T cells. Moreover, this B2M-/- hESC line contains no off-target integration or cleavage events, is devoid of stable B2M mRNA, exhibits a normal karyotype, and retains its self-renewal capacity, genomic stability, and pluripotency. Although B2M-/- hESC-derived cells are more susceptible to natural killer (NK) cells, murine transplantation studies have indicated that they are, overall, much less immunogenic than normal hESCs. Thus, these data show for the first time that, in vivo, the advantages provided by B2M-/- hESC-derived cells in avoiding CD8+ T-cell killing appear significantly greater than any disadvantage caused by increased susceptibility to NK cells.
Keywords: Differentiation and characterization; Human leukocyte antigen class I; Immunogenicity of human embryonic stem cells; β2-Microglobulin gene targeting strategy.
©AlphaMed Press.
Figures
References
-
- Thomson JA, Itskovitz-Eldor J, Shapiro SS, et al. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282:1145–1147. - PubMed
-
- Reubinoff BE, Pera MF, Fong CY, et al. Embryonic stem cell lines from human blastocysts: Somatic differentiation in vitro. Nat Biotechnol. 2000;18:399–404. - PubMed
-
- Mummery C, Ward-van Oostwaard D, Doevendans P, et al. Differentiation of human embryonic stem cells to cardiomyocytes: Role of coculture with visceral endoderm-like cells. Circulation. 2003;107:2733–2740. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
