

NH4(+) triggers the release of astrocytic lactate via mitochondrial pyruvate shunting
- PMID: 26286989
- PMCID: PMC4568276
- DOI: 10.1073/pnas.1508259112
NH4(+) triggers the release of astrocytic lactate via mitochondrial pyruvate shunting
Abstract
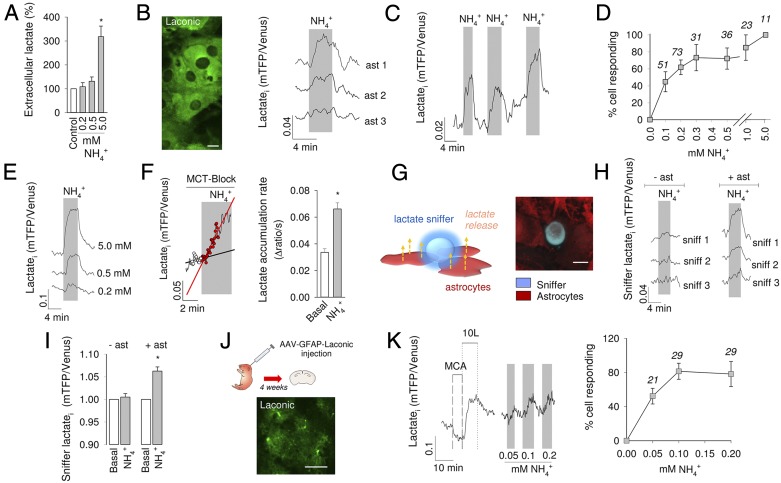
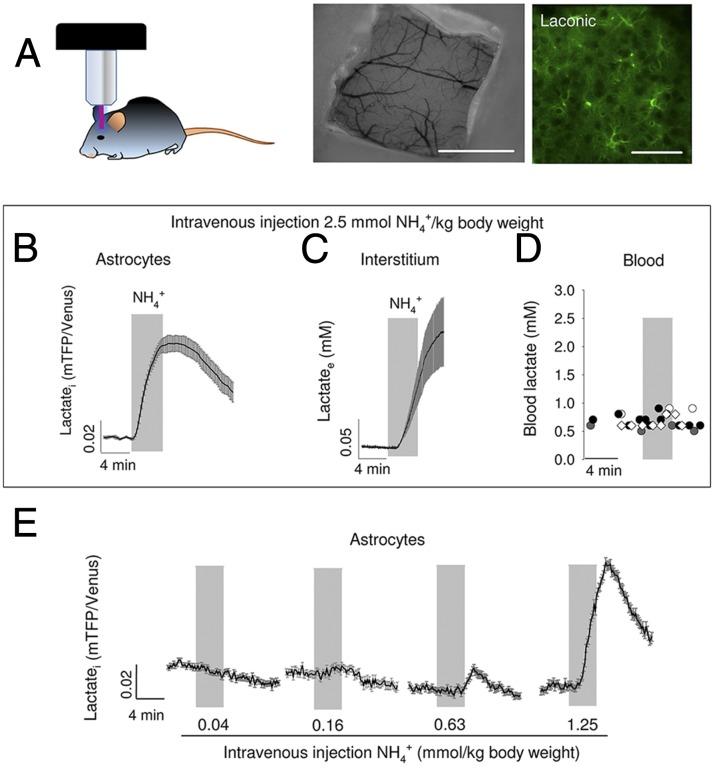
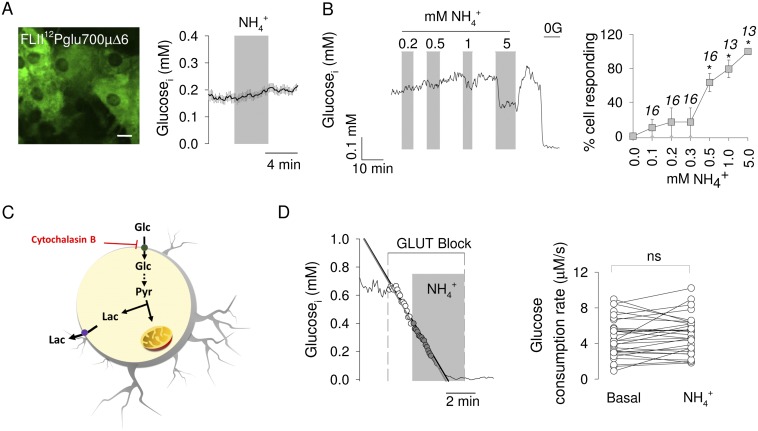
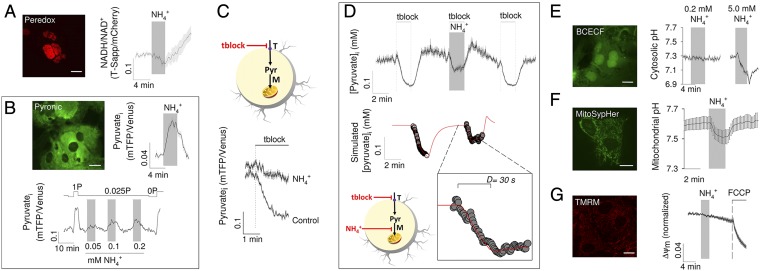
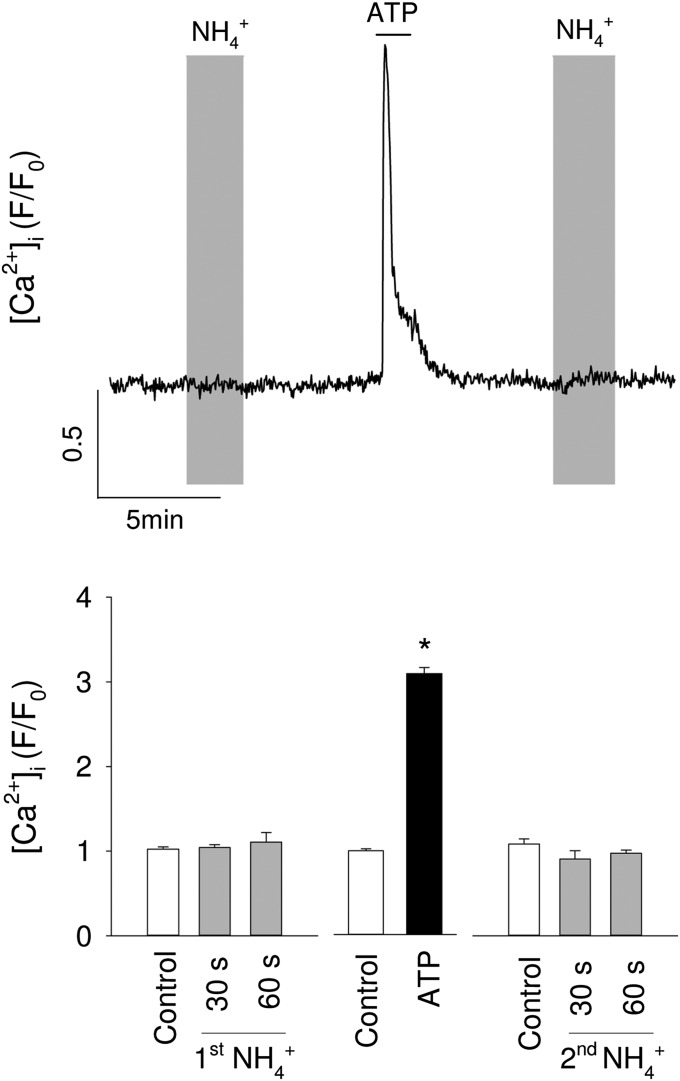
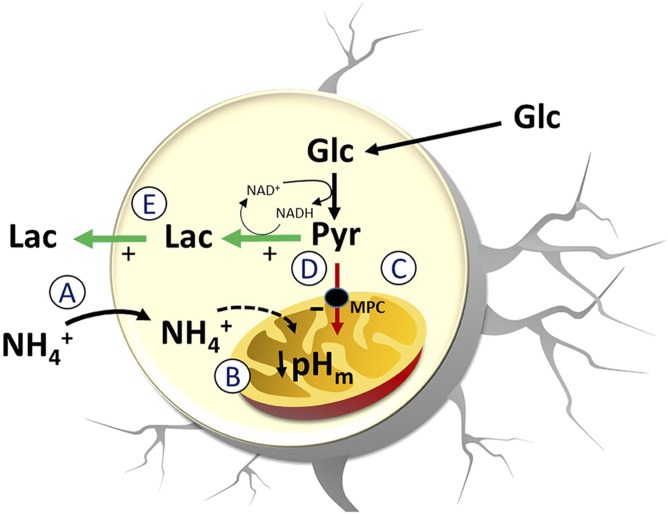
Neural activity is accompanied by a transient mismatch between local glucose and oxygen metabolism, a phenomenon of physiological and pathophysiological importance termed aerobic glycolysis. Previous studies have proposed glutamate and K(+) as the neuronal signals that trigger aerobic glycolysis in astrocytes. Here we used a panel of genetically encoded FRET sensors in vitro and in vivo to investigate the participation of NH4(+), a by-product of catabolism that is also released by active neurons. Astrocytes in mixed cortical cultures responded to physiological levels of NH4(+) with an acute rise in cytosolic lactate followed by lactate release into the extracellular space, as detected by a lactate-sniffer. An acute increase in astrocytic lactate was also observed in acute hippocampal slices exposed to NH4(+) and in the somatosensory cortex of anesthetized mice in response to i.v. NH4(+). Unexpectedly, NH4(+) had no effect on astrocytic glucose consumption. Parallel measurements showed simultaneous cytosolic pyruvate accumulation and NADH depletion, suggesting the involvement of mitochondria. An inhibitor-stop technique confirmed a strong inhibition of mitochondrial pyruvate uptake that can be explained by mitochondrial matrix acidification. These results show that physiological NH4(+) diverts the flux of pyruvate from mitochondria to lactate production and release. Considering that NH4(+) is produced stoichiometrically with glutamate during excitatory neurotransmission, we propose that NH4(+) behaves as an intercellular signal and that pyruvate shunting contributes to aerobic lactate production by astrocytes.
Keywords: FLII12Pglu700μΔ6; laconic; mitoSypHer; peredox; pyronic.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
