

Visual coding with a population of direction-selective neurons
- PMID: 26289471
- PMCID: PMC4620130
- DOI: 10.1152/jn.00919.2014
Visual coding with a population of direction-selective neurons
Abstract
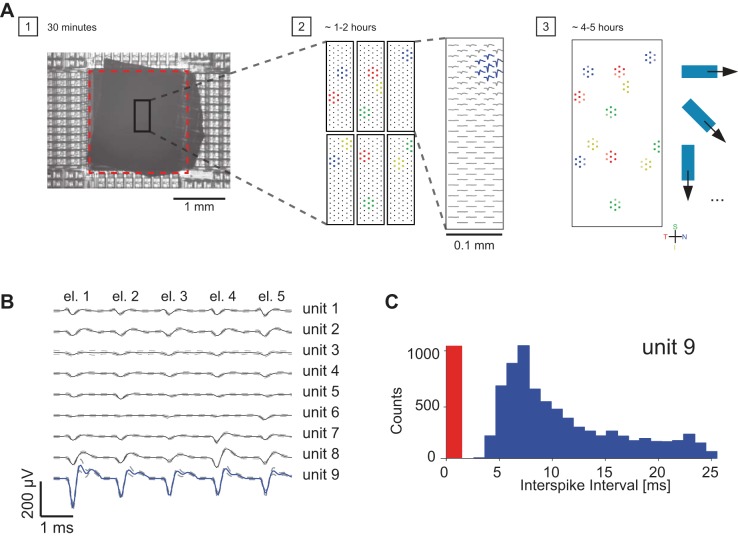
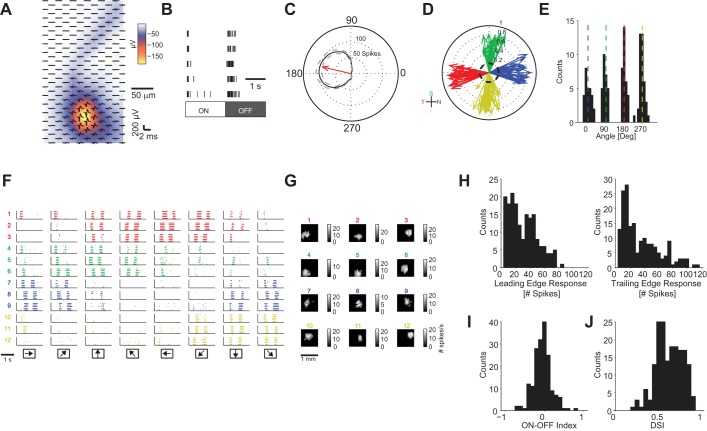
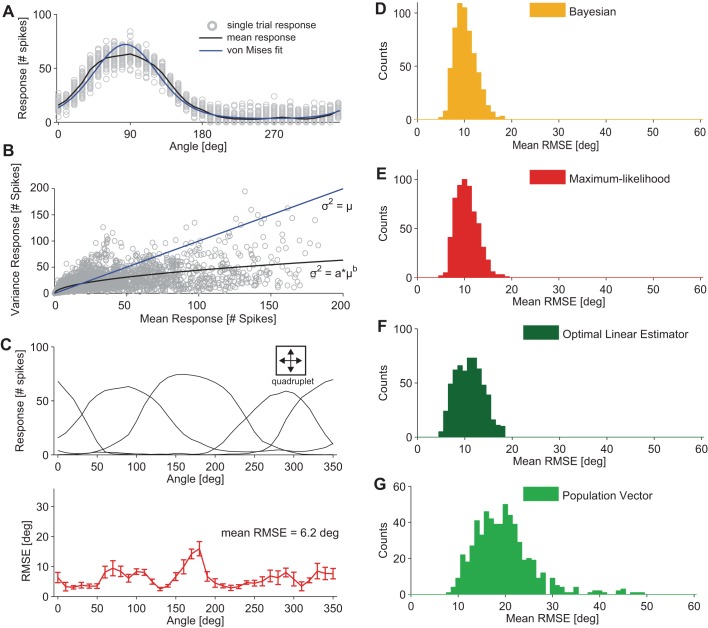
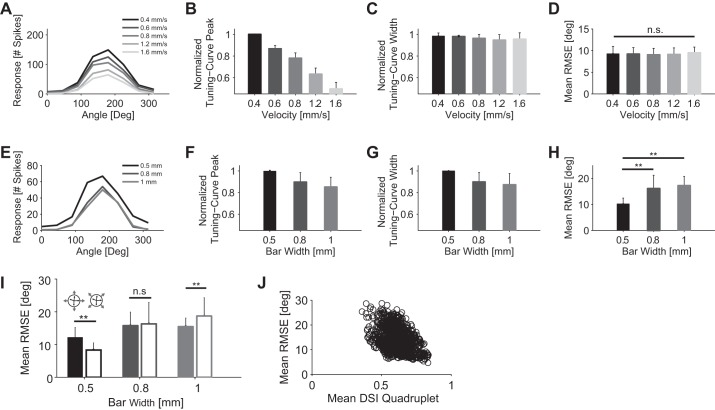
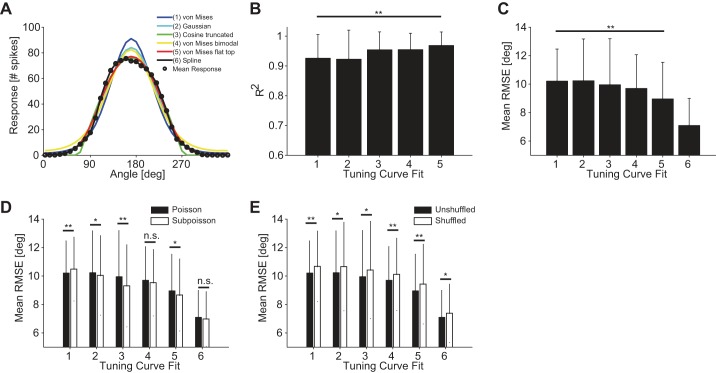
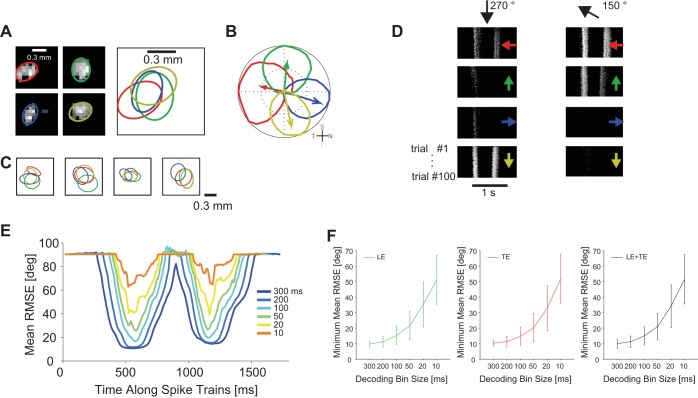
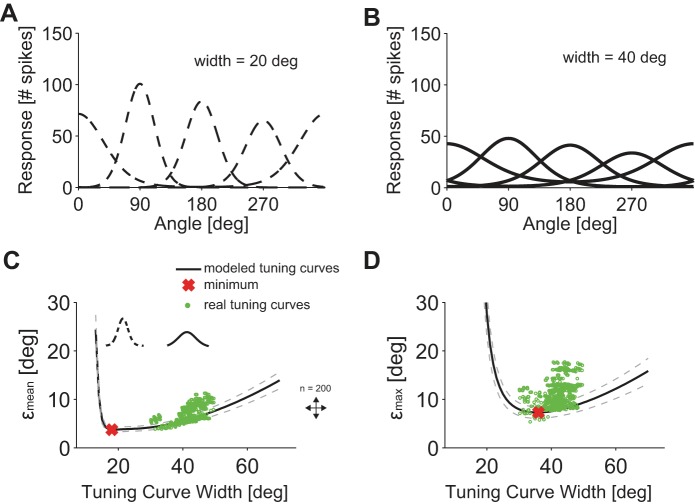
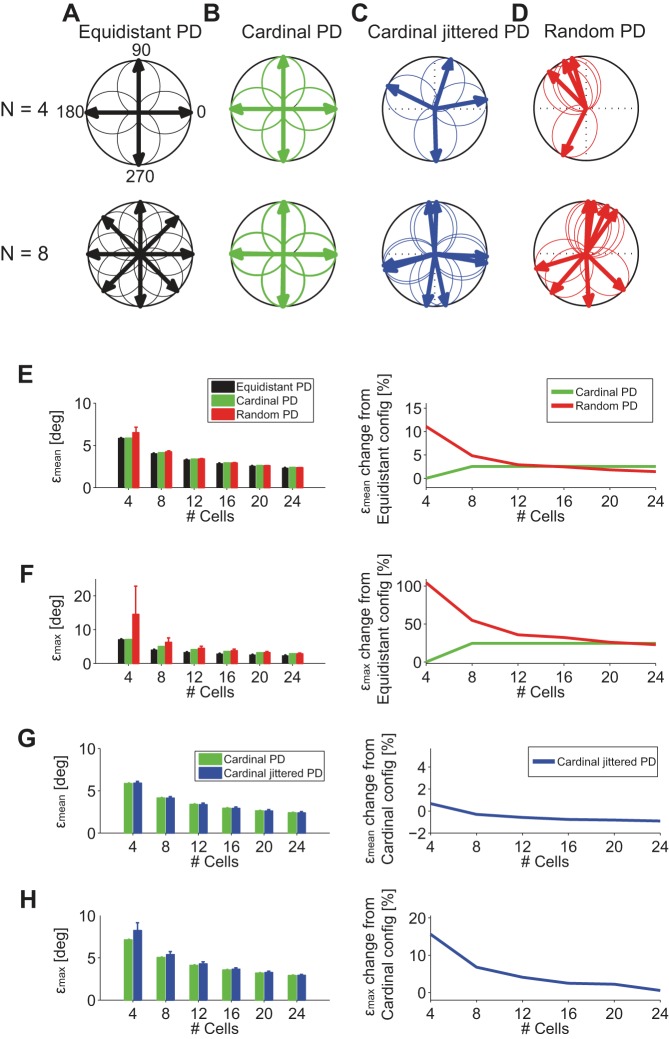
The brain decodes the visual scene from the action potentials of ∼20 retinal ganglion cell types. Among the retinal ganglion cells, direction-selective ganglion cells (DSGCs) encode motion direction. Several studies have focused on the encoding or decoding of motion direction by recording multiunit activity, mainly in the visual cortex. In this study, we simultaneously recorded from all four types of ON-OFF DSGCs of the rabbit retina using a microelectronics-based high-density microelectrode array (HDMEA) and decoded their concerted activity using probabilistic and linear decoders. Furthermore, we investigated how the modification of stimulus parameters (velocity, size, angle of moving object) and the use of different tuning curve fits influenced decoding precision. Finally, we simulated ON-OFF DSGC activity, based on real data, in order to understand how tuning curve widths and the angular distribution of the cells' preferred directions influence decoding performance. We found that probabilistic decoding strategies outperformed, on average, linear methods and that decoding precision was robust to changes in stimulus parameters such as velocity. The removal of noise correlations among cells, by random shuffling trials, caused a drop in decoding precision. Moreover, we found that tuning curves are broad in order to minimize large errors at the expense of a higher average error, and that the retinal direction-selective system would not substantially benefit, on average, from having more than four types of ON-OFF DSGCs or from a perfect alignment of the cells' preferred directions.
Keywords: coding; direction-selective system; microelectrode array; retina; retinal ganglion cells.
Copyright © 2015 the American Physiological Society.
Figures
References
-
- Amthor FR, Grzywacz NM, Merwine DK. Extra-receptive-field motion facilitation in on-off directionally selective ganglion cells of the rabbit retina. Vis Neurosci 13: 303–309, 1996. - PubMed
-
- Amthor FR, Oyster CW, Takahashi ES. Morphology of on-off direction-selective ganglion cells in the rabbit retina. Brain Res 298: 187–190, 1984. - PubMed
-
- Amthor FR, Takahashi ES, Oyster CW. Morphologies of rabbit retinal ganglion cells with complex receptive fields. J Comp Neurol 280: 97–121, 1989. - PubMed
-
- Amthor FR, Tootle JS, Grzywacz NM. Stimulus-dependent correlated firing in directionally selective retinal ganglion cells. Vis Neurosci 22: 769–787, 2005. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous