

Obesity-dependent changes in interstitial ECM mechanics promote breast tumorigenesis
- PMID: 26290412
- PMCID: PMC4837896
- DOI: 10.1126/scitranslmed.3010467
Obesity-dependent changes in interstitial ECM mechanics promote breast tumorigenesis
Abstract
Obesity and extracellular matrix (ECM) density are considered independent risk and prognostic factors for breast cancer. Whether they are functionally linked is uncertain. We investigated the hypothesis that obesity enhances local myofibroblast content in mammary adipose tissue and that these stromal changes increase malignant potential by enhancing interstitial ECM stiffness. Indeed, mammary fat of both diet- and genetically induced mouse models of obesity were enriched for myofibroblasts and stiffness-promoting ECM components. These differences were related to varied adipose stromal cell (ASC) characteristics because ASCs isolated from obese mice contained more myofibroblasts and deposited denser and stiffer ECMs relative to ASCs from lean control mice. Accordingly, decellularized matrices from obese ASCs stimulated mechanosignaling and thereby the malignant potential of breast cancer cells. Finally, the clinical relevance and translational potential of our findings were supported by analysis of patient specimens and the observation that caloric restriction in a mouse model reduces myofibroblast content in mammary fat. Collectively, these findings suggest that obesity-induced interstitial fibrosis promotes breast tumorigenesis by altering mammary ECM mechanics with important potential implications for anticancer therapies.
Copyright © 2015, American Association for the Advancement of Science.
Figures
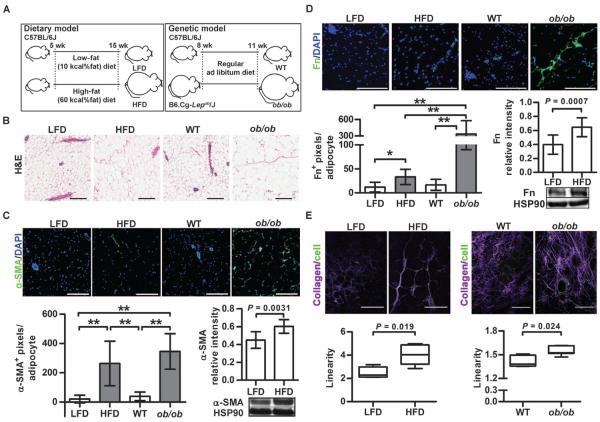
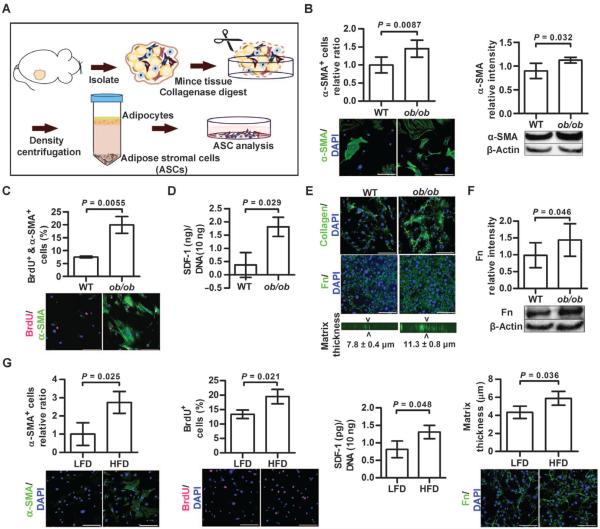
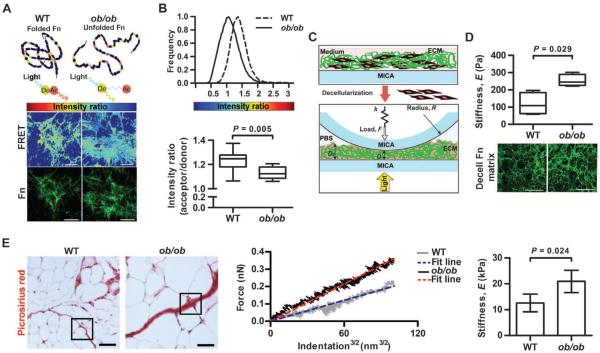
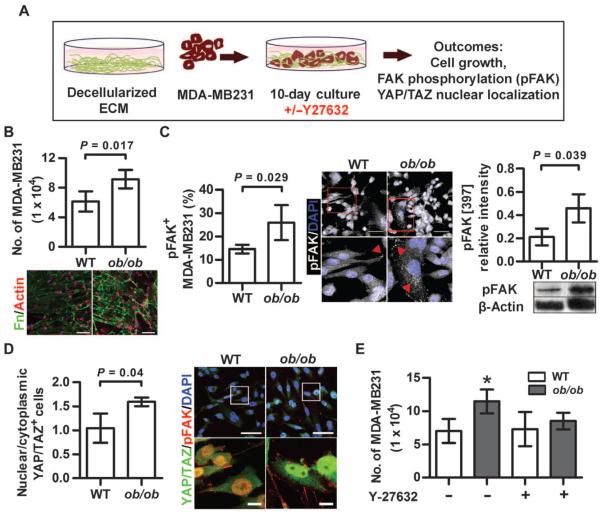
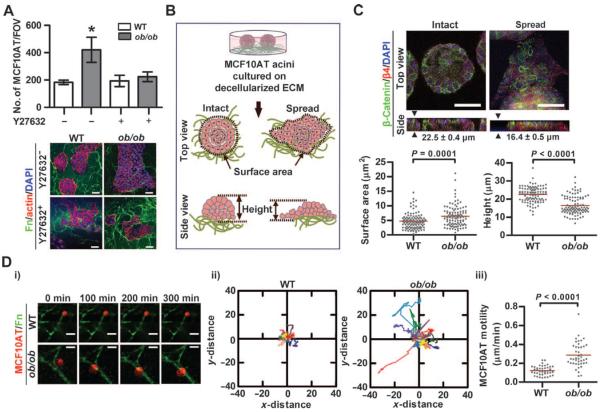
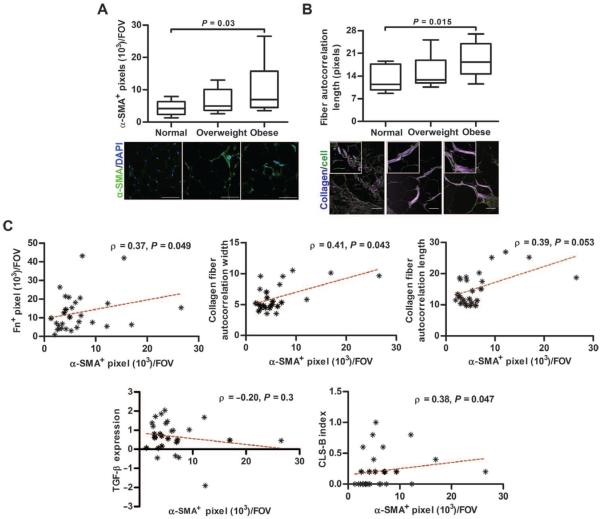
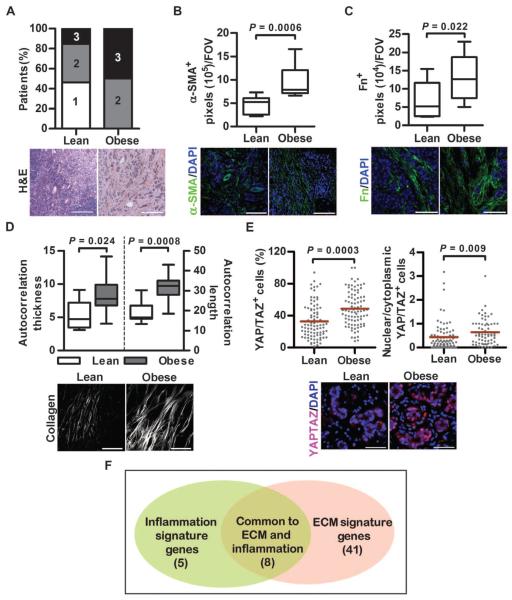
Comment in
-
Working stiff: how obesity boosts cancer risk.Sci Transl Med. 2015 Aug 19;7(301):301fs34. doi: 10.1126/scitranslmed.aac9446. Sci Transl Med. 2015. PMID: 26290410 Free PMC article.
-
Collagen and fibronectin: threads linking obesity and breast cancer.Ann Transl Med. 2016 Oct;4(Suppl 1):S50. doi: 10.21037/atm.2016.10.11. Ann Transl Med. 2016. PMID: 27868018 Free PMC article. No abstract available.
Similar articles
-
Obesity-Activated Adipose-Derived Stromal Cells Promote Breast Cancer Growth and Invasion.Neoplasia. 2018 Nov;20(11):1161-1174. doi: 10.1016/j.neo.2018.09.004. Epub 2018 Oct 12. Neoplasia. 2018. PMID: 30317122 Free PMC article.
-
CD44v6 increases gastric cancer malignant phenotype by modulating adipose stromal cell-mediated ECM remodeling.Integr Biol (Camb). 2018 Mar 1;10(3):145-158. doi: 10.1039/c7ib00179g. Epub 2018 Feb 16. Integr Biol (Camb). 2018. PMID: 29450424 Free PMC article.
-
Obesity-Associated Extracellular Matrix Remodeling Promotes a Macrophage Phenotype Similar to Tumor-Associated Macrophages.Am J Pathol. 2019 Oct;189(10):2019-2035. doi: 10.1016/j.ajpath.2019.06.005. Epub 2019 Jul 16. Am J Pathol. 2019. PMID: 31323189 Free PMC article.
-
Roles of obesity in the development and progression of breast cancer.Discov Med. 2017 Nov;24(133):183-190. Discov Med. 2017. PMID: 29278671 Review.
-
The effect of obesity on adipose-derived stromal cells and adipose tissue and their impact on cancer.Cancer Metastasis Rev. 2022 Sep;41(3):549-573. doi: 10.1007/s10555-022-10063-1. Epub 2022 Aug 24. Cancer Metastasis Rev. 2022. PMID: 35999486 Review.
Cited by
-
Cellular mechanisms linking cancers to obesity.Cell Stress. 2021 Apr 12;5(5):55-72. doi: 10.15698/cst2021.05.248. Cell Stress. 2021. PMID: 33987528 Free PMC article. Review.
-
Collagen microarchitecture mechanically controls myofibroblast differentiation.Proc Natl Acad Sci U S A. 2020 May 26;117(21):11387-11398. doi: 10.1073/pnas.1919394117. Epub 2020 May 8. Proc Natl Acad Sci U S A. 2020. PMID: 32385149 Free PMC article.
-
Stress Granules Determine the Development of Obesity-Associated Pancreatic Cancer.Cancer Discov. 2022 Aug 5;12(8):1984-2005. doi: 10.1158/2159-8290.CD-21-1672. Cancer Discov. 2022. PMID: 35674408 Free PMC article.
-
The covert symphony: cellular and molecular accomplices in breast cancer metastasis.Front Cell Dev Biol. 2023 Jun 27;11:1221784. doi: 10.3389/fcell.2023.1221784. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37440925 Free PMC article. Review.
-
Engineering the Extracellular Matrix to Model the Evolving Tumor Microenvironment.iScience. 2020 Oct 27;23(11):101742. doi: 10.1016/j.isci.2020.101742. eCollection 2020 Nov 20. iScience. 2020. PMID: 33225247 Free PMC article. Review.
References
-
- Khandekar MJ, Cohen P, Spiegelman BM. Molecular mechanisms of cancer development in obesity. Nat. Rev. Cancer. 2011;11:886–895. - PubMed
-
- Henegar C, Tordjman J, Achard V, Lacasa D, Cremer I, Guerre-Millo M, Poitou C, Basdevant A, Stich V, Viguerie N, Langin D, Bedossa P, Zucker JD, Clemen K. Adipose tissue transcriptomic signature highlights the pathological relevance of extracellular matrix in human obesity. Genome Biol. 2008;9:R14. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
- R01CA154481/CA/NCI NIH HHS/United States
- U54CA143876/CA/NCI NIH HHS/United States
- R01 CA185293/CA/NCI NIH HHS/United States
- R21CA161532/CA/NCI NIH HHS/United States
- 2UL1TR000457-06/TR/NCATS NIH HHS/United States
- P30 CA008748/CA/NCI NIH HHS/United States
- U54 CA143876/CA/NCI NIH HHS/United States
- 1S10RR025502/RR/NCRR NIH HHS/United States
- S10 RR025502/RR/NCRR NIH HHS/United States
- R21 CA161532/CA/NCI NIH HHS/United States
- R01 CA154481/CA/NCI NIH HHS/United States
- R01CA185293/CA/NCI NIH HHS/United States
- S10OD010605/OD/NIH HHS/United States
- UL1 TR000457/TR/NCATS NIH HHS/United States
- S10 OD010605/OD/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
