

Robust Anti-viral Immunity Requires Multiple Distinct T Cell-Dendritic Cell Interactions
- PMID: 26296422
- PMCID: PMC4567961
- DOI: 10.1016/j.cell.2015.08.004
Robust Anti-viral Immunity Requires Multiple Distinct T Cell-Dendritic Cell Interactions
Abstract
Host defense against viruses and intracellular parasites depends on effector CD8(+) T cells, whose optimal clonal expansion, differentiation, and memory properties require signals from CD4(+) T cells. Here, we addressed the role of dendritic cell (DC) subsets in initial activation of the two T cell types and their co-operation. Surprisingly, initial priming of CD4(+) and CD8(+) T cells was spatially segregated within the lymph node and occurred on different DCs with temporally distinct patterns of antigen presentation via MHCI versus MHCII molecules. DCs that co-present antigen via both MHC molecules were detected at a later stage; these XCR1(+) DCs are the critical platform involved in CD4(+) T cell augmentation of CD8(+) T cell responses. These findings delineate the complex choreography of cellular interactions underlying effective cell-mediated anti-viral responses, with implications for basic DC subset biology, as well as for translational application to the development of vaccines that evoke optimal T cell immunity.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
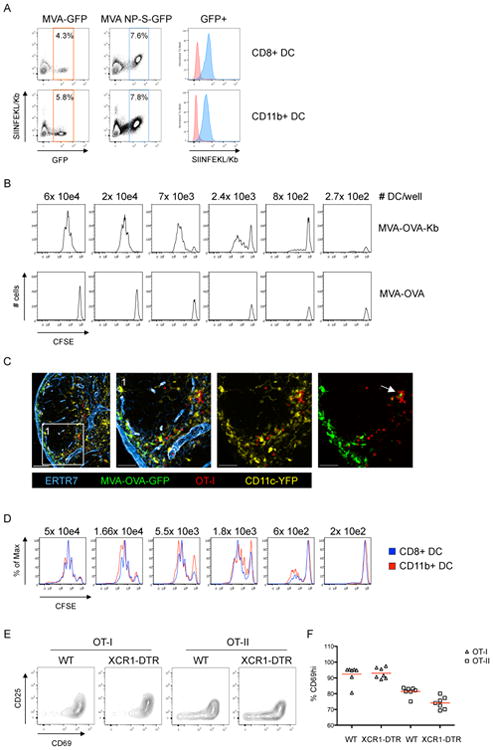
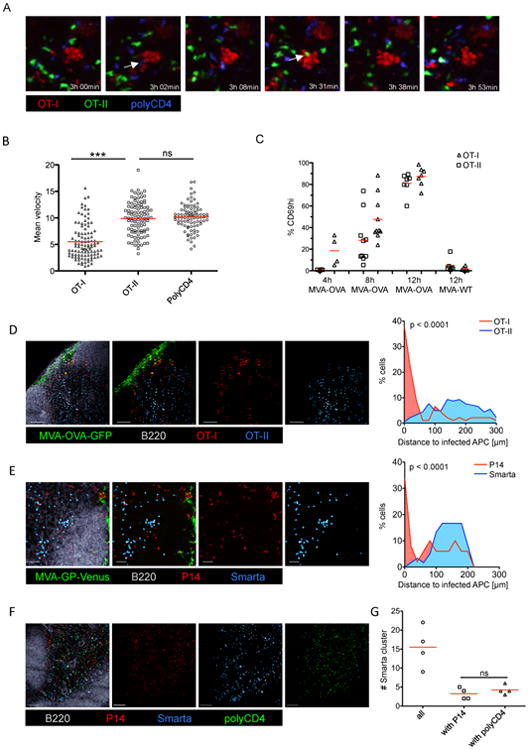
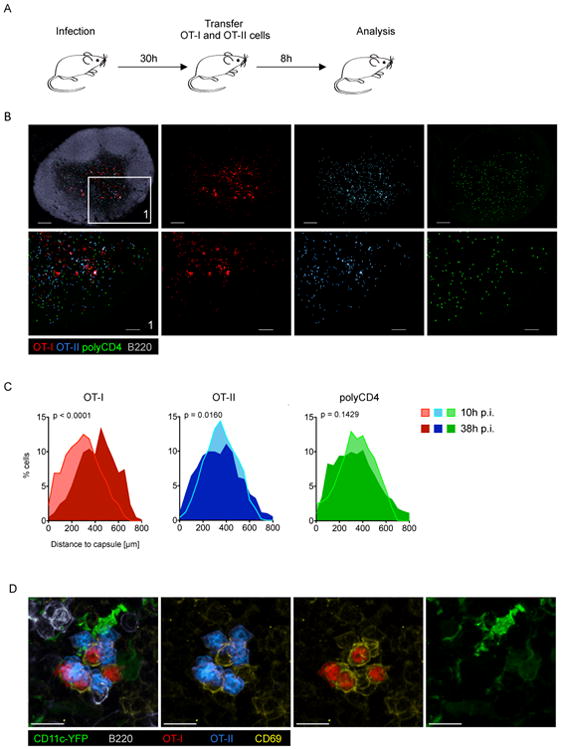
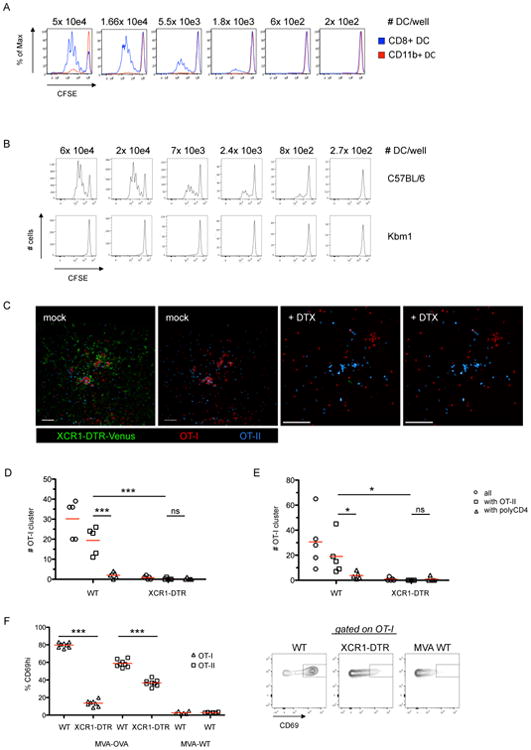
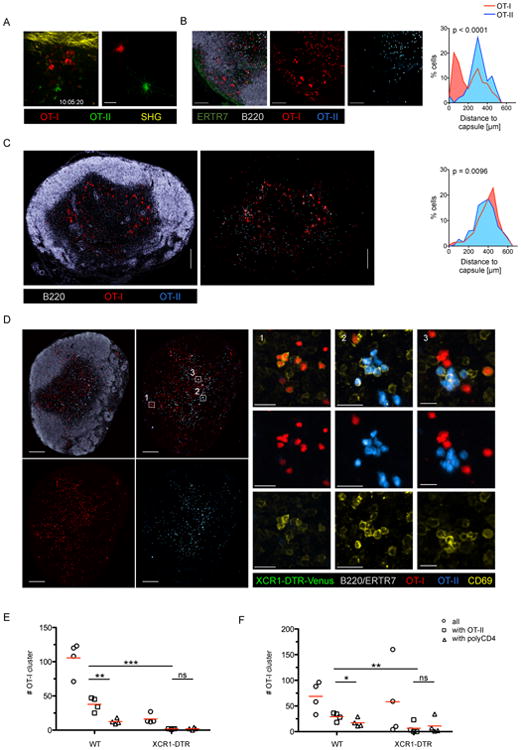
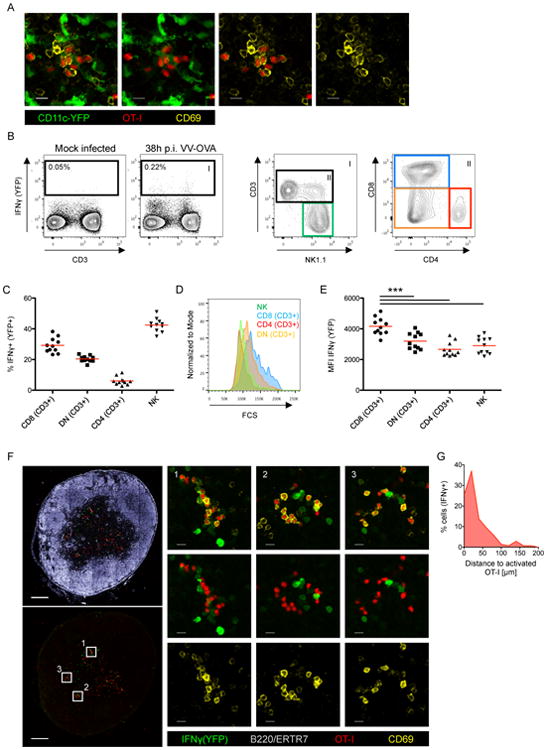
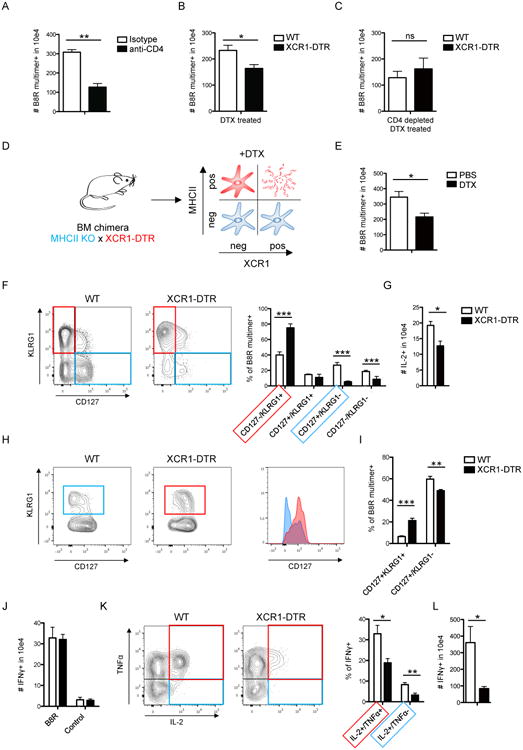
Comment in
-
Helping the Help for CD8+ T Cell Responses.Cell. 2015 Sep 10;162(6):1210-2. doi: 10.1016/j.cell.2015.08.051. Cell. 2015. PMID: 26359981
References
-
- Allan RS, Waithman J, Bedoui S, Jones CM, Villadangos JA, Zhan Y, Lew AM, Shortman K, Heath WR, Carbone FR. Migratory dendritic cells transfer antigen to a lymph node-resident dendritic cell population for efficient CTL priming. Immunity. 2006;25:153–162. - PubMed
-
- Becker M, Guttler S, Bachem A, Hartung E, Mora A, Jakel A, Hutloff A, Henn V, Mages HW, Gurka S, et al. Ontogenic, Phenotypic, and Functional Characterization of XCR1(+) Dendritic Cells Leads to a Consistent Classification of Intestinal Dendritic Cells Based on the Expression of XCR1 and SIRPalpha. Front Immunol. 2014;5:326. - PMC - PubMed
-
- Bedoui S, Whitney PG, Waithman J, Eidsmo L, Wakim L, Caminschi I, Allan RS, Wojtasiak M, Shortman K, Carbone FR, et al. Cross-presentation of viral and self antigens by skin-derived CD103+ dendritic cells. Nat Immunol. 2009;10:488–495. - PubMed
-
- Bennett SR, Carbone FR, Karamalis F, Flavell RA, Miller JF, Heath WR. Help for cytotoxic-T-cell responses is mediated by CD40 signalling. Nature. 1998;393:478–480. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
