

Protein kinase C-related kinase 1 and 2 play an essential role in thromboxane-mediated neoplastic responses in prostate cancer
- PMID: 26296974
- PMCID: PMC4694913
- DOI: 10.18632/oncotarget.4664
Protein kinase C-related kinase 1 and 2 play an essential role in thromboxane-mediated neoplastic responses in prostate cancer
Abstract
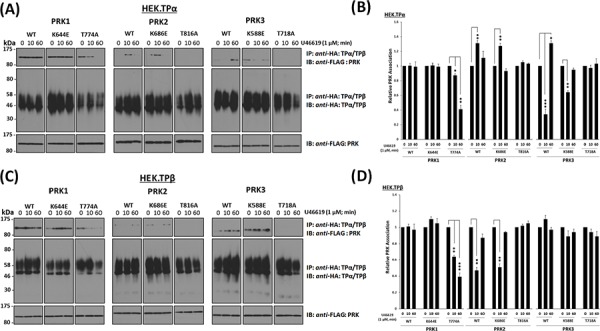
The prostanoid thromboxane (TX) A2 is increasingly implicated in neoplastic progression, including prostate cancer (PCa). Mechanistically, we recently identified protein kinase C-related kinase (PRK) 1 as a functional interactant of both the TPα and TPβ isoforms of the human T prostanoid receptor (TP). The interaction with PRK1 was not only essential for TPα/TPβ-induced PCa cell migration but also enabled the TXA2-TP axis to induce phosphorylation of histone H3 at Thr11 (H3Thr11), an epigenetic marker both essential for and previously exclusively associated with androgen-induced chromatin remodelling and transcriptional activation. PRK1 is a member of a subfamily of three structurally related kinases comprising PRK1/PKNα, PRK2/PKNγ and PRK3/PKNβ that are widely yet differentially implicated in various cancers. Hence, focusing on the setting of prostate cancer, this study investigated whether TPα and/or TPβ might also complex with PRK2 and PRK3 to regulate their activity and neoplastic responses. While TPα and TPβ were found in immune complexes with PRK1, PRK2 and PRK3 to regulate their activation and signalling, they do so differentially and in a TP agonist-regulated manner dependent on the T-loop activation status of the PRKs but independent of their kinase activity. Furthermore, TXA2-mediated neoplastic responses in prostate adenocarcinoma PC-3 cells, including histone H3Thr11 phosphorylation, was found to occur through a PRK1- and PRK2-, but not PRK3-, dependent mechanism. Collectively, these data suggest that TXA2 acts as both a neoplastic and epigenetic regulator and provides a mechanistic explanation, at least in part, for the prophylactic benefits of Aspirin in reducing the risk of certain cancers.
Keywords: cancer; prostate; protein kinase C-related kinase; receptor; thromboxane.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures







References
-
- Choi J, Kermode JC. New therapeutic approaches to combat arterial thrombosis: better drugs for old targets, novel targets, and future prospects. Mol Interv. 2011;11:111–123. - PubMed
-
- Davi G, Santilli F, Vazzana N. Thromboxane receptors antagonists and/or synthase inhibitors. Handb Exp Pharmacol. 2012:261–286. - PubMed
-
- Zha S, Yegnasubramanian V, Nelson WG, Isaacs WB, De Marzo AM. Cyclooxygenases in cancer: progress and perspective. Cancer Lett. 2004;215:1–20. - PubMed
-
- Rothwell PM, Wilson M, Elwin CE, Norrving B, Algra A, Warlow CP, Meade TW. Long-term effect of aspirin on colorectal cancer incidence and mortality: 20-year follow-up of five randomised trials. Lancet. 2010;376:1741–1750. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
