

Neural Population Evidence of Functional Heterogeneity along the CA3 Transverse Axis: Pattern Completion versus Pattern Separation
- PMID: 26298276
- PMCID: PMC4548827
- DOI: 10.1016/j.neuron.2015.07.012
Neural Population Evidence of Functional Heterogeneity along the CA3 Transverse Axis: Pattern Completion versus Pattern Separation
Abstract
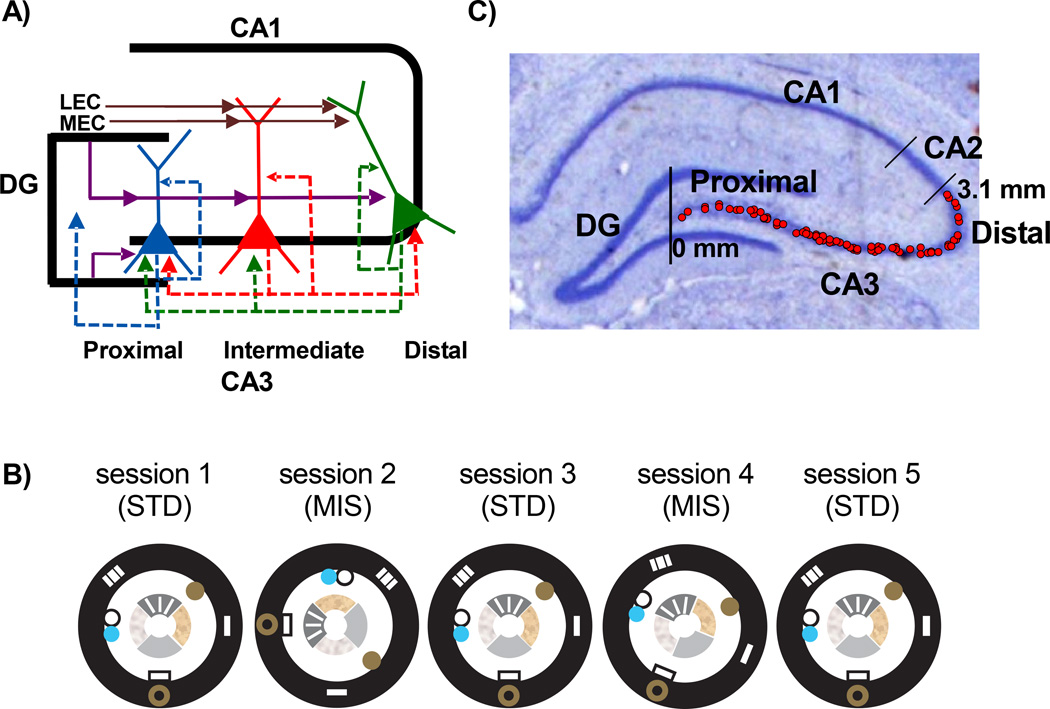
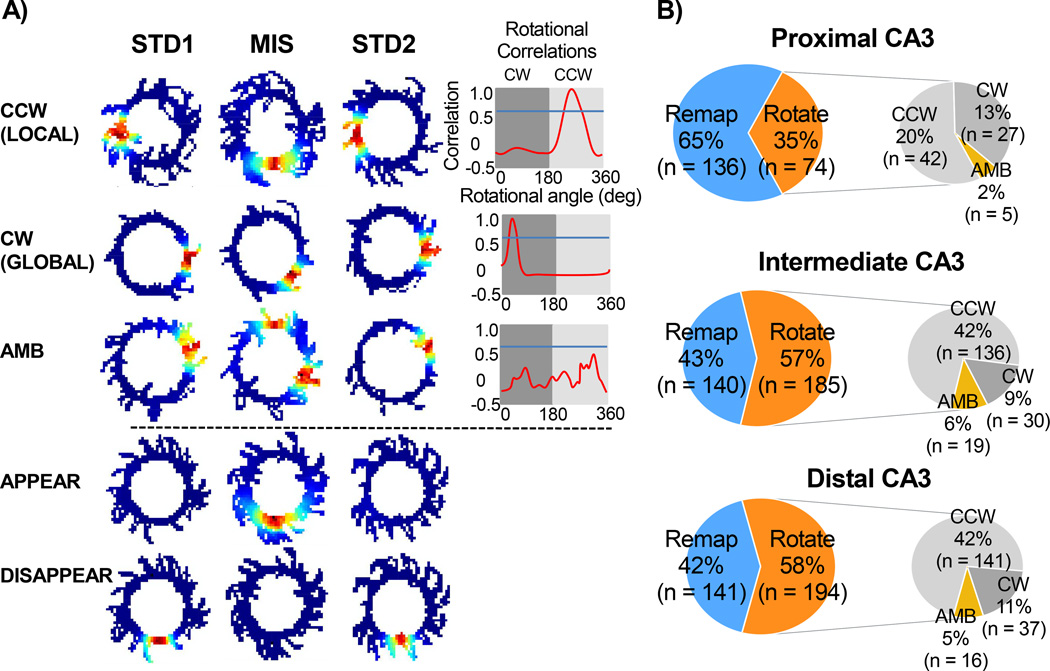
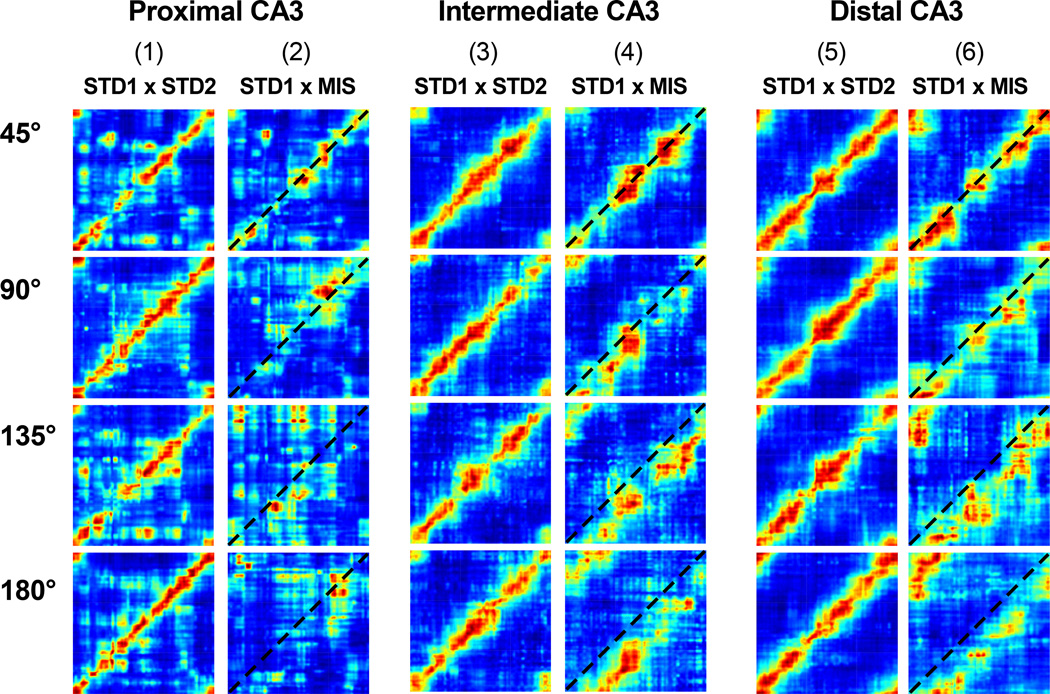
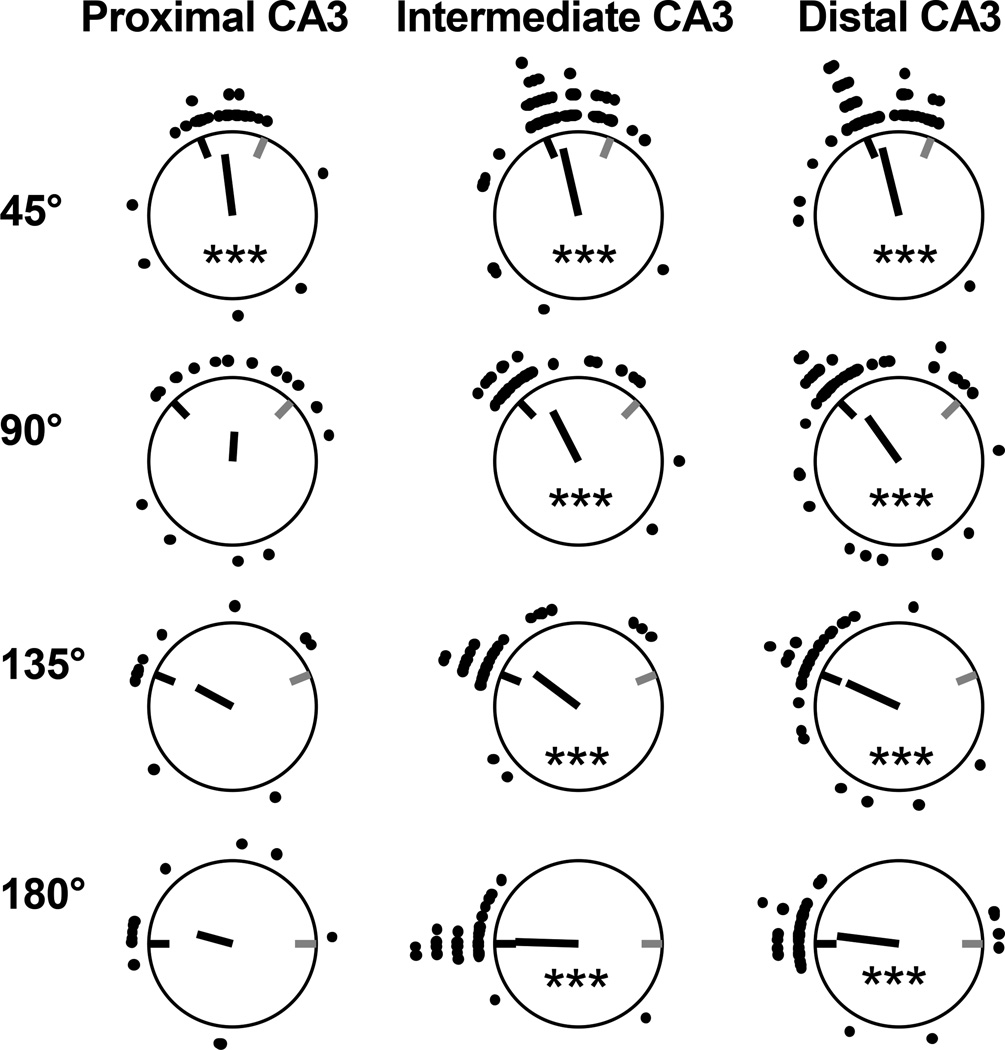
Classical theories of associative memory model CA3 as a homogeneous attractor network because of its strong recurrent circuitry. However, anatomical gradients suggest a functional diversity along the CA3 transverse axis. We examined the neural population coherence along this axis, when the local and global spatial reference frames were put in conflict with each other. Proximal CA3 (near the dentate gyrus), where the recurrent collaterals are the weakest, showed degraded representations, similar to the pattern separation shown by the dentate gyrus. Distal CA3 (near CA2), where the recurrent collaterals are the strongest, maintained coherent representations in the conflict situation, resembling the classic attractor network system. CA2 also maintained coherent representations. This dissociation between proximal and distal CA3 provides strong evidence that the recurrent collateral system underlies the associative network functions of CA3, with a separate role of proximal CA3 in pattern separation.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures




Comment in
-
Spatial processing: mapping function.Nat Rev Neurosci. 2015 Oct;16(10):575. doi: 10.1038/nrn4029. Epub 2015 Sep 9. Nat Rev Neurosci. 2015. PMID: 26350242 No abstract available.
References
-
- Amaral DG, Witter MP. The three-dimensional organization of the hippocampal formation: a review of anatomical data. Neuroscience. 1989;31:571–591. - PubMed
-
- Amit DJ. Modeling Brain Function: The World of Attractor Neural Networks. Cambridge, UK/New York: Cambridge Univ. Press; 1989.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
