

Curcumin as therapeutics for the treatment of head and neck squamous cell carcinoma by activating SIRT1
- PMID: 26299580
- PMCID: PMC4547100
- DOI: 10.1038/srep13429
Curcumin as therapeutics for the treatment of head and neck squamous cell carcinoma by activating SIRT1
Abstract
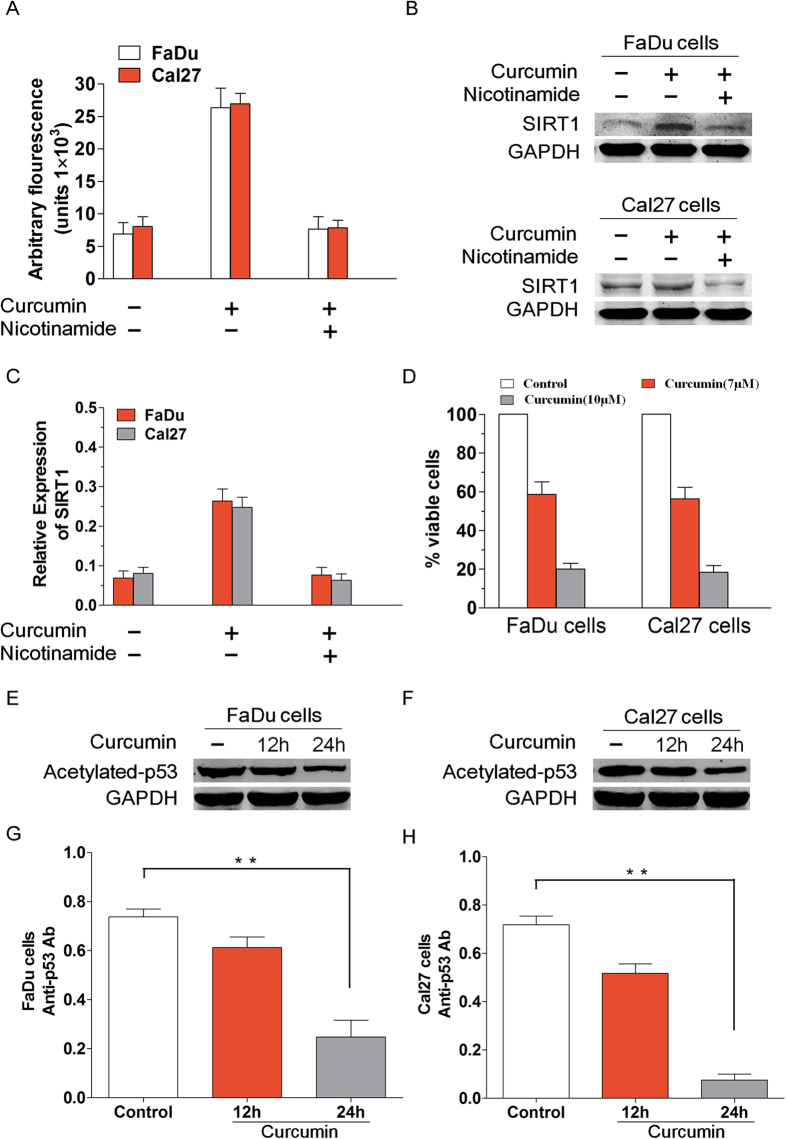
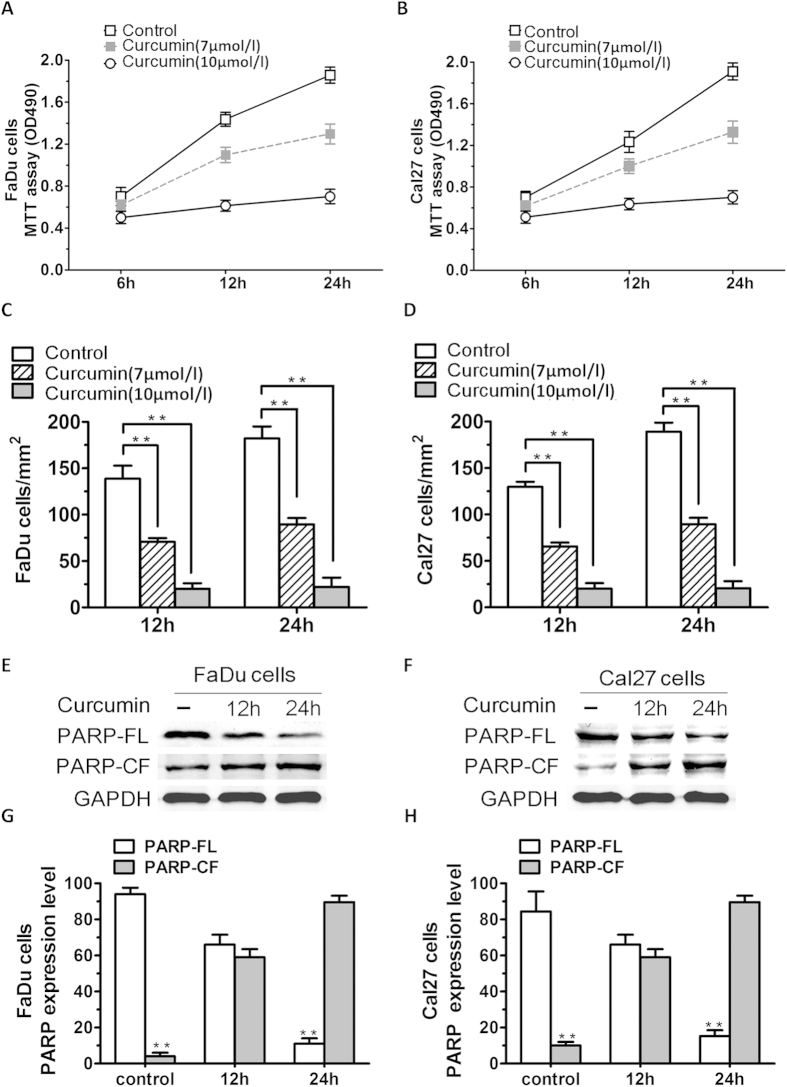
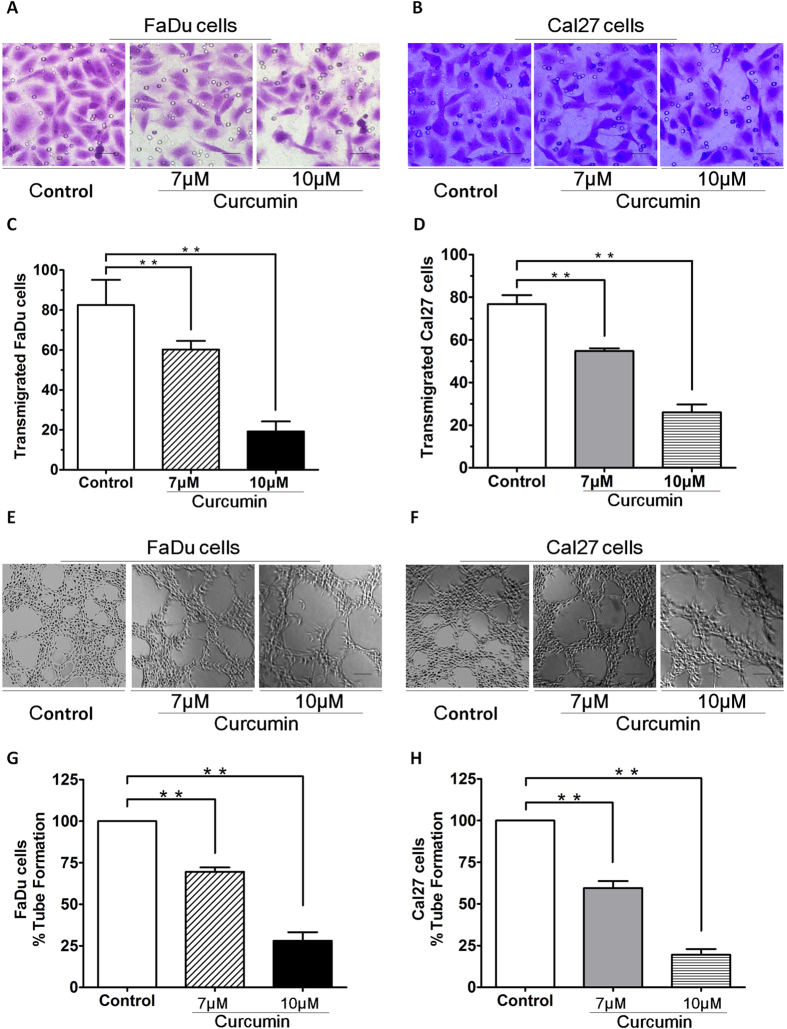
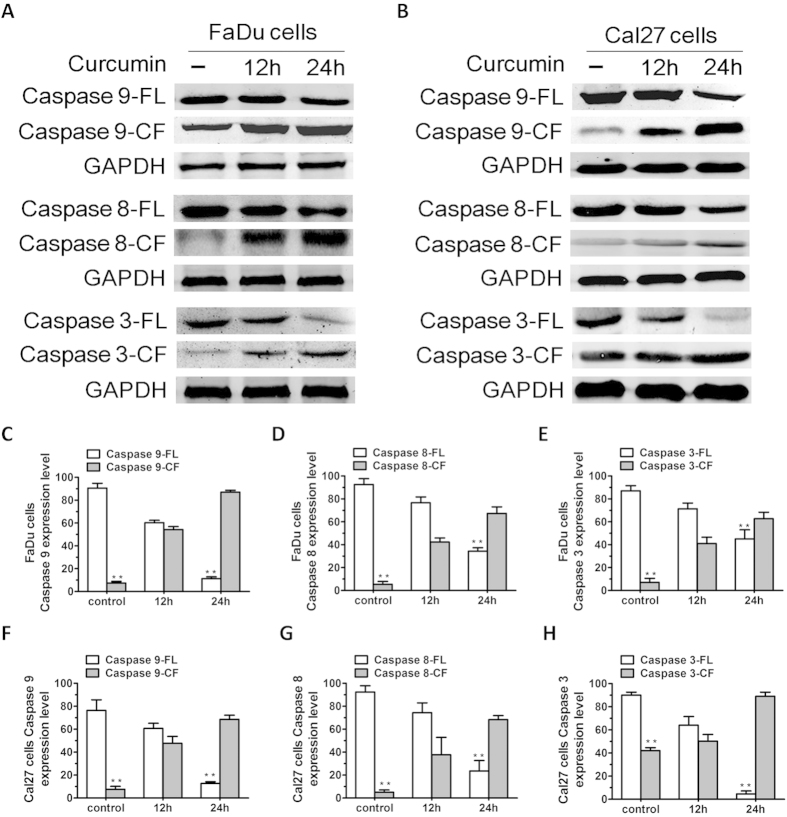
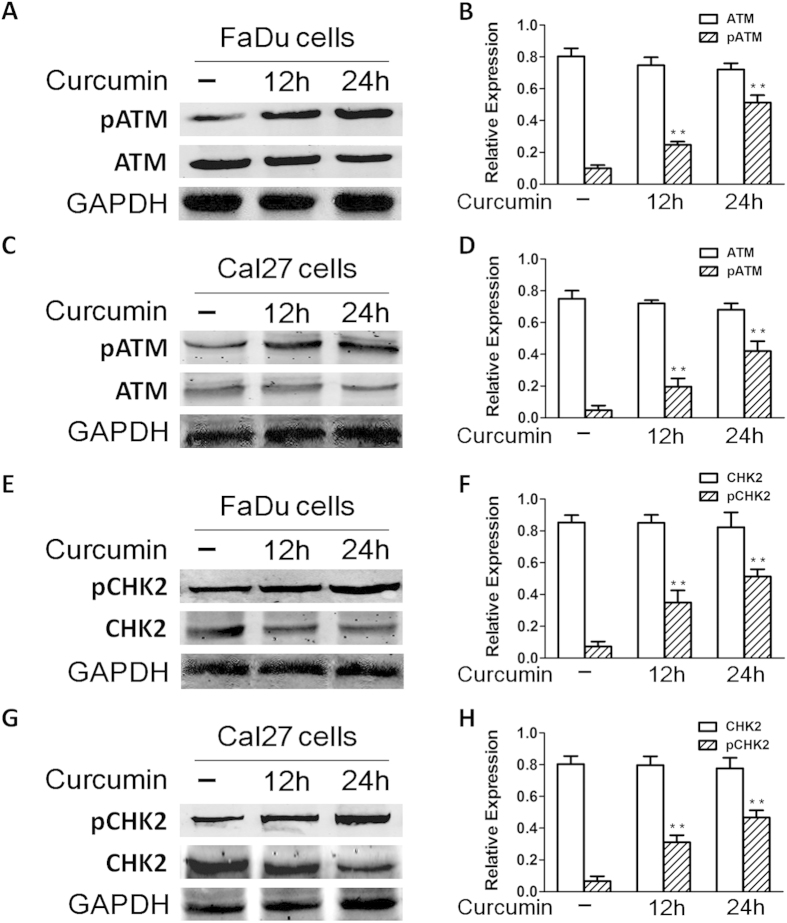
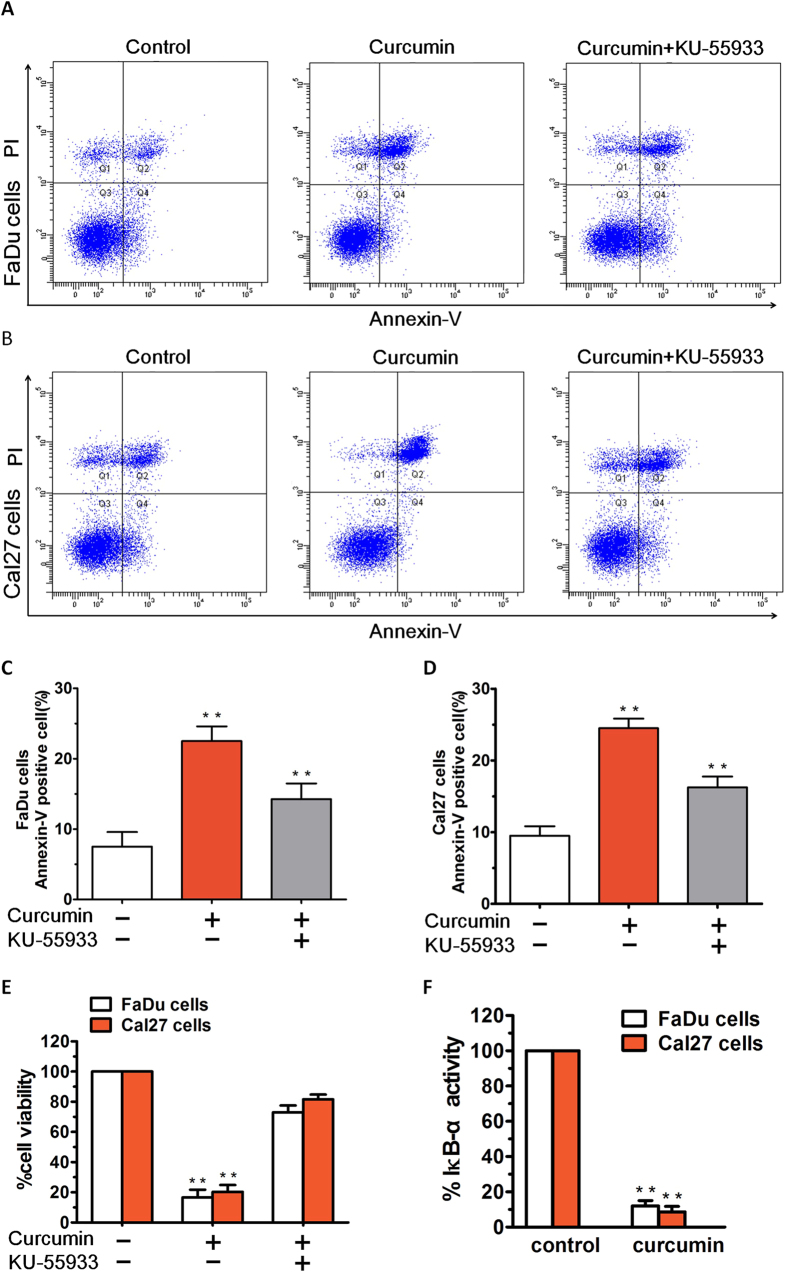
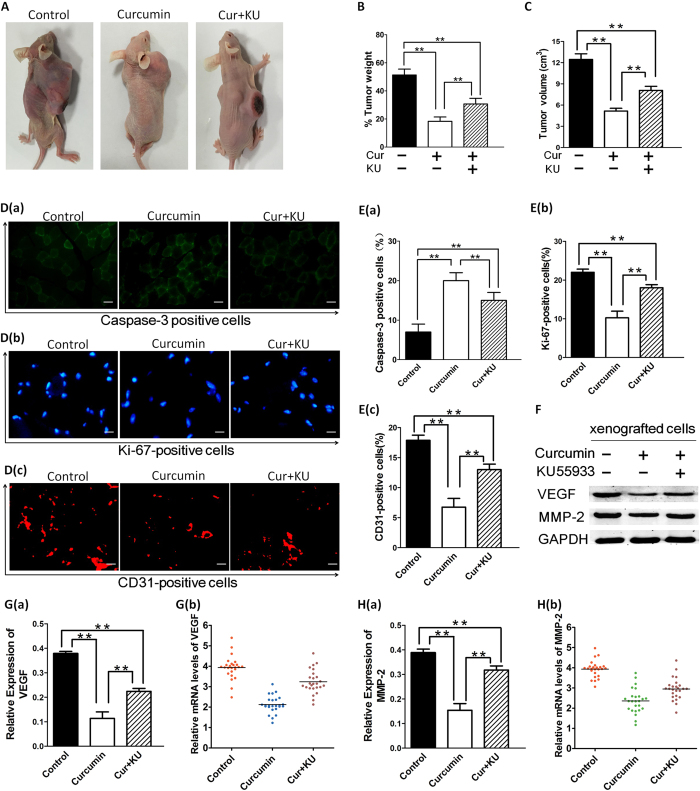
SIRT1 is one of seven mammalian homologs of Sir2 that catalyzes NAD(+)-dependent protein deacetylation. The aim of the present study is to explore the effect of SIRT1 small molecule activator on the anticancer activity and the underlying mechanism. We examined the anticancer activity of a novel oral agent, curcumin, which is the principal active ingredient of the traditional Chinese herb Curcuma Longa. Treatment of FaDu and Cal27 cells with curcumin inhibited growth and induced apoptosis. Mechanistic studies showed that anticancer activity of curcumin is associated with decrease in migration of HNSCC and associated angiogenesis through activating of intrinsic apoptotic pathway (caspase-9) and extrinsic apoptotic pathway (caspase-8). Our data demonstrating that anticancer activity of curcumin is linked to the activation of the ATM/CHK2 pathway and the inhibition of nuclear factor-κB. Finally, increasing SIRT1 through small molecule activator curcumin has shown beneficial effects in xenograft mouse model, indicating that SIRT1 may represent an attractive therapeutic target. Our studies provide the preclinical rationale for novel therapeutics targeting SIRT1 in HNSCC.
Figures
References
-
- He L. et al. Mcl-1 and FBW7 control a dominant survival pathway underlying HDAC and Bcl-2 inhibitor synergy in squamous cell carcinoma. Cancer Discov 3, 324–337, 10.1158/2159-8290.cd-12-0417 (2013). - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
